

Motboken avskaffades samma år som jag föddes. Som ett ödets ironi har jag nu suttit här, trots min frus suckanden, i några veckor och lusläst och kommenterat docent Lars Johan Erkells (i fortsättningen rätt och slätt E.) bok Intelligent Design – vetenskap, gudsbevis eller politisk agenda (Universus Academic Press, 2025). Det hela skulle visa sig bli en hel motbok. E. noterade också i sitt förord att även hans fru haft en liknande inställning som min. En eloge till dem båda för att de uthärdat allt tangentknatter utan hysteriska utbrott.
Så här presenteras boken på Bokus hemsida:
Var kommer vi ifrån? Är livet ett resultat av blinda naturlagar - eller finns det ett syfte, en plan, en design bakom allt? Ja, svarar teorin om intelligent design. Den erbjuder ett till synes vetenskapligt alternativ till evolutionen. Teorin är en del i det kulturkrig mellan liberal demokrati och religiös fundamentalism som just nu utspelas i USA, och som har återverkningar även i Sverige.I denna populärvetenskapliga debattbok visar E. med skärpa, kunskap och berättarglädje hur teorin vuxit fram, inte ur nya vetenskapliga rön, utan som en strategi för att återinföra religiösa idéer i klassrummet och samhället.
Boken synar noga argumenten för intelligent design och visar varför de inte håller. En gedigen genomgång av vad vetenskap är och inte är, liksom en saklig och balanserad diskussion av förhållandet mellan religion och vetenskap gör boken lämplig för alla - inte minst för studiegrupper - som intresserar sig för förhållandet mellan vetenskap och religion.
En tankeväckande, provocerande och aktuell bok om kunskapens roll i samhället - och varför det spelar roll vad vi bygger vår världsbild på.
Det är jag som fetstilat de tre formuleringarna. Speciellt de båda första kommer att vara i fokus i den här recensionen.
Lars Johan presenterade själv sin bok i humanisternas monter under Bokmässan i Göteborg i höstas (2025).
Och jag hade glädjen att få samtala med honom om en del påståenden i boken i Genesispodden 2026-03-04.
I tabellen här nedanför hittar du en förteckning över bokens olika kapitel. De är länkade till mina kommentarer så att du lätt kan få en nyanserad bild av det kapitel du just läst. Detta ifall du har tillgång till boken vill säga, men jag har ansträngt mig för att du ska kunna läsa min motbok även utan boken till hands. Om du klickar på en viss not så kommer du att länkas till notförklaringen i slutet av dokumentet. Du återvänder via den lilla symbolen ”↩”.
Föredrar du att skriva ut dokumentet och läsa i pappersform så kan du klicka här eller på PDF-symbolen nedan. Då får du sidnoter i stället för slutnoter nederst på varje sida och länkarna även i form av kortlänkar som är lätta att skriva in i en sökmotor vid behov. Men dokumentet är på lite drygt hundra sidor, så kör dubbelsidigt för miljöns skull, och se till att ha papper i skrivaren 😉.

Klicka på ikonen för att läsa eller skriva ut som PDF
Givande läsning tillönskas!
Göran
Bokens struktur
Citat i dokumentet som inte är Lars Johan Erkells kursiveras i den fortsatta texten
Inledning
Det var med stort intresse blandat med vissa farhågor1 som jag tog mig an den här boken. Mina farhågor besannades under läsningen. När jag dessutom insåg att boken högst sannolikt kommer att finnas på bibliotek och institutioner i skolor och på universitet, inte minst på de teologiska institutionerna som redan nu är avogt inställda till evolutionskritik, så insåg jag motvilligt att jag inte hade något val: Det måste helt enkelt finnas en tillgänglig ”motbok” till E.s kraftigt vinklade redogörelse av intelligent design-rörelsen (i fortsättningen ID) och dess argument. Nu är vi där.
Risk för missförstånd
En läsare som inte är insatt i evolutionsbiologi och inte heller är väl orienterad om hur man tänker och resonerar inom modern kreationism och ID-rörelse kommer förmodligen att bli övertygad av E.s resonemang. Han skriver till synes sakligt, strukturerat och genomtänkt och med en hel del referenser till forskningens frontlinjer. Och med en lagom dos av sarkasmer mot hur han anser att man resonerar inom ID-rörelsen.
Läsaren kommer att uppfatta evolutionsteorin som den enda objektiva och relevanta ursprungsförklaringen baserad på all tillgänglig evidens av experiment och observationer. ID-perspektivet kommer att framstå som ett religiöst pseudovetenskapligt och rent antivetenskapligt önsketänkande och som en central del i ett politiskt-religiöst program för att tvinga världens befolkning till ideologisk underkastelse – alla måste bli kristna och följa Gamla testamentets lagar och bud (kap 29). Typ så.
Den som så småningom har tagit del av det jag nu ska förmedla kommer emellertid att inse att när E.s budskap i den här boken analyserats och satt i perspektiv och de flesta av hans argument förångats så är det inte så mycket som återstår i botten av kastrullen. Men det lilla som finns kvar där är absolut värt att fundera över. Jag kommer att förklara närmare i sinom tid.
När man som E. är docent i biologi vid ett statligt universitet åtnjuter man ett avsevärt mått av respekt och trovärdighet när man formulerar sig, kanske speciellt hos kategorier av människor som till vardags befinner sig på ett visst avstånd från naturvetenskapen. Naturvetenskapen har ju fortfarande en ganska hög status i samhället, eftersom den i mångas ögon ses som bärare av den objektiva sanningen om verkligheten. Med respekten följer ett ansvar. Om man beskyller andra för ovetenskaplighet följer ett desto större ansvar för att det man påstår är vetenskapligt välgrundat.
Jag vill betona att jag inte har något som helst skäl att hävda att E. skulle vara vare sig okunnig, oärlig eller ha några dolda avsikter. Han är uppriktigt övertygad om evolutionens faktum och följaktligen lika övertygad om att ID-rörelsen är fel ute. Han är också oroad för konsekvenserna av ett scenario där den religiösa högern med ID som murbräcka ska skapa oönskade samhällsförändringar på båda sidorna av Atlanten.
Men hur goda intentioner man än har så kan det bli fel. Och när sanningar, halvsanningar och felaktigheter blandas och presenteras med retorisk skicklighet, då och då i kombination med historierevisionism, måste varningsflaggan hissas.
Inte helt poänglös
Detta till trots vill jag som sagt redan här i inledningen betona att E.s bok har en och annan poäng. Jag håller till exempel med honom om att Intelligent design-rörelsen har brister när det gäller teoribygget. Det är faktiskt lättare att formulera teoribaserade prediktioner utifrån biblisk kreationism än från ID.
Och det alltid nyttigt för alla att få mothugg. Jag är övertygad om den saken, och ska man ta E. på orden så gäller det även honom. Han skriver:
Det ständiga ifrågasättandet och den öppna debatten är det bästa skyddet mot ortodoxi och dogmatism. (s 65)
Det påminner om bibelordet:
Järn skärper järn, den ena människan den andra. (Ord 27:17)
Det är viktigt att höra båda parter innan man tar ställning, i det här fallet bedömer ID-rörelsens argument. Min förhoppning är därför att var och en som läser E.s bok också ska läsa den här motboken. Och faktiskt även vice versa, trots de brister det ska visa sig att den har.
Det här kan säkert uppfattas som en gnällig inledning från min sida. Men jag håller med E.:
Om nu någon skulle tycka att jag är väl kritisk, vill jag påpeka att det är så den vetenskapliga debatten förs. (s 83)
Om man ger sig in i leken får man helt enkelt leken tåla.
![]()
Låt oss från början ställa oss frågan: Vad är egentligen vetenskap? En klassisk filosofisk definition är Sokrates (469-399 f Kr) princip att följa spåren varthän de leder, eller annorlunda formulerat: att söka sanningen om verkligheten oavsett hur den ser ut.
Wikipedia definierar vetenskap så här: Vetenskap är produktionen av ny kunskap med systematiska metoder.
Ytterligare en definition är: Aktiviteter där man genom att utveckla teorier söker att förklara observationer i naturen.
E. är obekväm med den sortens definitioner. Han tycker att det saknas något. Vad kan det vara? Vi återkommer strax till det (för den som är alltför nyfiken - se punkt 5 nedan).
![]()
AVDELNING I
Boken första avdelning som omfattar kapitlen 1-10 har rubriken Vetenskap. I de första sju kapitlen presenterar E. ett antal grundläggande vetenskapsteoretiska begrepp. Det är här E. lägger sin idémässiga grund för att förkasta ID och evolutionskritiken.
I förordet nämner E. att han under flera år varit medlem av en dialoggrupp tillsammans med ett antal religiösa personer som han också namnger i (s 8), däribland några präster i Svenska kyrkan (se not 57). Det hade varit intressant att ta del av deras synpunkter på den här motboken.
↩ Tillbaka till innehållsförteckningen
![]()
Prologen kan sammanfattas i dess sista två meningar:
Det är bättre att erkänna sin okunnighet än att hitta på en förklaring. Då slipper man lura sig själv. Och – ännu viktigare – man lurar inte andra. (s 11)
E. har rätt. Men som vi ska se kommer samma formulering att kunna användas mot honom själv.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 1. Om vetenskapliga förklaringar
Jag har ingenting att kommentera i det här kapitlet, förutom att formuleringen
Därför har förklaringar baserade på gudar eller demoner ingen förklaringskraft; de är påståenden vi bara griper ur luften. (s 16)
kommer att vara ett återkommande mantra boken igenom. E. anser att ingenting i världen skulle kunna indikera förekomsten av en skapare.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 2. Faktum, lag, hypotes och teori
Jag vill redan nu flagga för att jag kommer att avsätta gott om utrymme för att analysera och ge perspektiv på det E. behandlar här, eftersom det ligger till grund för den kritik han sedan riktar mot skapelse-/designperspektivet i bokens huvuddel (kap 11-25).
De viktigaste sakerna som framhålls är:
1. Evolutionsmodellen uppfyller kriterierna för att få kallas en teori, och det gör den immun mot kritik:
En vetenskaplig teori skall säga något om varför världen ser ut som den gör, och inte på något annat sätt. Det innebär att den samtidigt förbjuder fenomen som skulle strida mot teorin. (s 20)
2. E. anser att de båda vetenskapliga arbetssätten induktion (generella slutsatser utifrån begränsade observationer, s 23) liksom den hypotetisk-deduktiva metoden (formulerande och prövande av hypoteser, s 24) bekräftar evolutionsteorin. ID däremot, menar han, bygger på abduktion (s 24, slutledning till den bästa förklaringen), en metod som dominerar inom historiska vetenskaper och som utgår från indicier (indirekta spår) i stället för direkta observationer. E. konstaterar att denna metod är mindre tillförlitlig än de båda andra såvida den inte underbyggs med ”solid evidens”. (s 24)
3. Andra metoder som Ekell avråder från är analogibevis (s 34, t ex likheterna mellan ett öga och en kamera), intuition (s 34, samt kap 24 s 157, verkligheten kan ibland vara annorlunda än man tror) och bevis från okunnigheten (s 35, när man utgår från vad man inte vet i stället för från det man vet).
4. Reduktionism förekommer i två former: dels i form av en grundläggande metod där man först studerar de enskilda delarna i ett system med syftet att sedan kunna förstå helheten. Och dels i form av ett grundläggande antagande att alla biologiska fenomen kan förklaras av underliggande kemiska och fysikaliska reaktioner och principer. (s 27)
5. Vetenskapligt arbete måste enligt E. utgå från en metodologisk naturalism, 2 (s 38) som till sin natur är agnostisk. Naturalism förekommer även som en ateistisk filosofi.3 (s 39) E. menar att ID-förespråkare vantolkar den metodologiska naturalismen som om den vore filosofisk naturalism. (s 39)
Eftersom naturalismen är en av vetenskapens grundprinciper kommer enligt E. varje hänvisning till en icke-materiell designer att per definition betraktas som ovetenskaplig. Därmed elimineras den vetenskapsteoretiska grunden för intelligent design, eftersom både designern och vederbörandes skapelsemetod ligger utanför naturalismens ram. (s 45)
6. E. erkänner att det inte finns några allmängiltiga demarkationskriterier (regler för att åtskilja vetenskap från ovetenskap). (s 46) Därefter presenterar han likväl ett antal sådana kriterier som han tycker är lämpliga: falsifierbarhet (s 47, att det ska vara möjligt att motbevisa ens teori), relevanta metoder för arbetet (s 49), saklig och relevant argumentation (s 49) samt etablering (s 50, att forskningen bedrivs vid seriösa institutioner av välutbildade forskare och att resultaten referentgranskas innan publicering.
Mina kommentarer till bokens första avdelning
Uppenbarligen har E. har två syften med den här inledande avdelningen: Dels att betona evolutionsteorins ställning som objektiv vetenskap och att all evidens understryker att den är så nära ett historiskt faktum man kan komma. Dels att inleda arbetet med att underminera ett antal argument som brukar lyftas fram av företrädare för ID-rörelsen som stöd för att designmodellen är vetenskaplig.
Jag kommer nu att ge mina reflektioner till E.s sju punkter parallellt med genomgången av de fem följande bokkapitlen.
1. Evolutionsteorins status som teori. (s 20)
Den dominerande uppfattningen bland nutida biologer – och många andra – är att evolutionen är mer än en teori, den är ett ”faktum”.4 Det råder inget tvivel om att E. tillhör denna kategori.
Det är förmodligen därför som han inte uppmärksammar det ironiska i att han ägnar en hel bok åt att argumentera för att evolutionskritiker har fel och evolutionsbiologer rätt, samtidigt som han redan i inledningen konstaterar att evolutionsteorin uttryckligen förbjuder alla argument som skulle kunna anföras mot den. Vad finns det då för anledning att argumentera?
E. menar säkert inte det här. Jag tror att han uttryckte sig slarvigt och egentligen avsåg begreppet ”naturlag”. En naturlag förbjuder fenomen som strider mot den, men en teori gör det naturligtvis inte.
Jag vill påstå att den egentliga orsaken till E.s tvärsäkerhet är av filosofisk natur. Den motiveras inte av evolutionsargumenten som sådana – de är, som vi snart ska se häpnadsväckande bristfälliga – utan av en annan slags övertygelse.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 3. Om naturvetenskapligt arbete
2. De vetenskapliga arbetssätten.
E. tar ett konkret exempel när han vill visa att evolutionsteorin stöds via induktionsmetoden, nämligen likheten mellan människor och schimpanser (s 23), som ju närmast är en ikon för evolutionen.
I en bok som behandlar ID-rörelsen kunde han ha nämnt att man inom denna rörelse menar att likheter mellan organismer kan ha andra förklaringar är biologiskt släktskap. Ibland är det naturligtvis så, som de mellan oss och våra barn, eller mellan olika slags änder, men i andra fall tolkar man likheterna helt olika beroende på vilken grundsyn man har på ursprungsfrågan. Kreationister anser att hierarkiska mönster av likheter och skillnader är en designmarkör, det vill säga något som avspeglar en medveten aktörs skaparstrategi snarare än spår efter ett evolutionärt släktskap. E. har här, i likhet med de flesta evolutionister, en blind fläck i synfältet. Man har mycket svårt att förstå att likheter mellan organismer i den levande världen kan ha andra förklaringar är biologiskt släktskap.
Abduktion
E. vet om att ID-förespråkare som Stephen Meyer och William Dembski argumenterar för ID:s vetenskaplighet just genom hänvisning till att design är den bästa av tillgängliga förklaringar det vill säga de för ett abduktivt resonemang.
Jag tolkar E.s strategi här som följer:
a. Det går inte att förneka att det dominerande arbetssättet inom historiska vetenskaper är via abduktion till den bästa förklaringen.
b. Det går inte heller att förneka att evolutionsteorin till stora delar är en historisk vetenskap och att de evolutionära scenarion som målas upp till stor del bygger på indicier och inte på direkta observationer.5
c. Men att betrakta evolutionsteorin som en historisk vetenskap ger ID-anhängarna ett vetenskapsteoretiskt alibi att hävda att designteori kvalificerar sig som vetenskap.
d. Följaktligen är det säkrast att hävda att det är irrelevant att dela upp vetenskapen i historisk respektive experimentell vetenskap.6 (s 36)
De historiska delarna av evolutionsteorin är med nödvändighet mindre tillförlitliga. Eller åtminstone inte tillförlitligare än den evidens man har tillgång till.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 4. Om bevis och evidens i vetenskapen
3. Argument som E. avråder ifrån.
Det mest klassiska och intuitiva av alla argument för Guds existens är analogiargumentet (s 32). I sitt klassiska verk Natural Theology or Evidences of the Existence and Attributes of the Deity från 1802 argumenterar den brittiske prästen och filosofen William Paley (1743-1805) 7 för att företeelser i naturen vittnar om sin skapare på ett snarlikt sätt som mänskligt designade föremål vittnar om sina formgivare. Det mest berömda exempel som Paley nämner i sin bok är när en vandrare upptäcker ett fick-ur på en hed och drar slutsatsen att klockans uppenbara tecken på design pekar mot dess designer i form av en urmakare. Den analogin är också bakgrunden till rubriken på ateisten Richard Dawkins välkända bok Den blinde urmakaren (The Blind Watchmaker) från 1986, där denne argumenterar för att någon urmakare inte behövs i naturen eftersom det naturliga urvalet räcker gott och väl.
E. argumenterar för att alla analogier mellan företeelser i naturen och mänskliga konstruktioner är felaktiga jämförelser (s 32-34). Varför då? Vi återkommer till det.
Vid upprepade tillfällen anklagar E. ID-förespråkare för att argumentera från okunnigheten. Jag kommer även att behandla det när det blir aktuellt.
4. Reduktionism. (s 26)
Som metod är ett reduktionistiskt arbetssätt helt okontroversiellt. Det är helt enkelt så man brukar arbeta när man ska undersöka någon komplex företeelse. Man studerar en del i sänder och använder sedan resultaten till att förstå hur helheten fungerar.
Men den form av reduktionism som inte alls är lika självklar är den filosofiska variant som E. antyder att han också anser vara relevant. Det är att varje fenomen i naturen ytterst kan förklaras av sina underliggande nivåer, vilket är liktydigt med ett förnekande av att helheten skulle vara någonting förmer än alla delarna. Varje biologisk struktur eller funktion skulle i så fall kunna reduceras till kemi, och ytterst till fördelningen av elektriska laddningar hos atomer och molekyler (atomfysik).
Ett annat uttryck för samma idé är tron att alla komplexa strukturer i den levande världen utgör resultatet av en ofantligt lång kedja av ackumulerade (plussade) små obetydliga förändringar.
Min gissning är att det är därför som E. attraheras av tanken, eller åtminstone lämnar fältet öppet för att det skulle kunna förhålla sig så. Det
är ingenting man kan veta … det är ett antagande man gör (s 27).
Vid flera tillfällen längre fram i boken visar han att han själv är anhängare av filosofisk reduktionism.8 Det torde inte råda något tvivel om att om någon valt att hålla sig till en sådan syn så får det återverkningar inom många områden av ens tänkande och världsbild.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 5. Naturalism, vad är det?
Kapitel 6. Naturliga och övernaturliga förklaringar
Jag kommenterar de här båda kapitlen tillsammans eftersom de i princip förmedlar samma budskap.
Jag nämnde i inledningen att E. tyckte att det saknades något i de där generella definitionerna av vetenskap. Det är naturalismen. Det finns några centrala argument som han lyfter fram i boken. Det här är ett av dem: Eftersom ID-hypotesen för fram existensen av en Designer som inte ryms inom den materiella verkligheten så är ID per definition ovetenskaplig. Vetenskapen förbjuder inblandning av icke-materiella storheter, den måste ovillkorligt arbeta under premissen av metodologisk naturalism (s 29, 38).
Nu finns det ingenting som säger att verkligheten med nödvändighet skulle låta sig inordnas i något visst system som vi människor konstruerat. Vår uppgift inom vetenskapen borde i stället vara att skapa ramar för vår forskning som fångar in så stor del av verkligheten som möjligt – eller om man så vill, sanningen om den.
Med den begränsning som E. förordar skulle forskningen aldrig komma åt sanningen, utan bara strängt materialistiska uttryck för den. De frågeställningar forskarna ställer kommer aldrig att kunna vara: ”Vilken är den egentliga orsaken till fenomenet X?” utan ”Vilka är de materialistiska orsakerna till fenomenet X? Enda omständigheten när det inte skulle spela någon roll vilken av frågorna man ställer är om det bara vore materialistiska förklaringar som existerade. Men det går inte att veta. E. formulerar det så här:
Man kan inte studera övernaturliga makter man inte har någon kunskap om. (s 45)
Vi återkommer till den utsagan när vi diskuterar ”vetenskapsargumentet” lite senare (kap 12), men redan nu vill jag bemöta E.s resonemang med att påstå att vi inte behöver veta hur någonting har skett och vem som gjort något för att kunna slå fast att det har skett och att någon har gjort det. För det sistnämnda behöver vi inte lämna den naturalistiska principen en enda sekund. Inom forensisk vetenskap, som till exempel brottsutredningar, är den första frågan man ställer sig huruvida det som inträffat är föranlett av en ren tillfällighet eller om det skett med en bakomliggande avsikt. Om man då kommer fram till att det verkligen finns en avsikt (brottslig handling), först då börjar arbetet med att identifiera gärningsmannen. ID fokuserar främst på den första frågan, helt inom ramen för vedertagna naturalistiska principer.
E. och jag är överens om att man måste skilja mellan naturalismen som metod och filosofi. Men vad spelar det för roll när så många andra underlåtit det under åren och tillåtits sprida sina vanföreställningar?
Uttalanden som det av George Gaylord Simpson, dåtidens ledande neo-darwinist har påverkat generationer:
Människan är resultatet av en planlös och materialistisk process som inte hade henne i åtanke. Hon var inte planerad.9
Andra som hävdat att darwinismen gjort en materialistisk världsbild mer trovärdig, och likaså att materialistiska evolutionära teorier gjort religiösa trosföreställningar om Gud antingen ohållbara eller mindre trovärdiga, är Douglas Futuyma, William Provine, Stephen Jay Gould och Richard Dawkins. Dawkins sade ju som många vet att:
Darwin gjorde det möjligt att bli en intellektuellt tillfredsställd ateist.
Det faktum att somliga ateistiska naturvetare överbetonat sin metodologiska naturalism till att bli metafysisk har inte på något vis hindrat darwinismens status som vetenskaplig teori. Vilken rätt har man då att påstå att ID som vetenskaplig teori måste avfärdas som ovetenskaplig på grund av att kristna forskare har en teistisk syn på världen? Det är naturligtvis en teoris kvalifikationer och inte dess eventuella implikationer som ska avgöra dess relevans.
↩ Tillbaka till slutet av introduktionen
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 7. Vetenskap och pseudovetenskap
6. Demarkationskriterier. Först konstaterar E. att det inte finns några generella kriterier för att särskilja vetenskap från pseudovetenskap (s 46). Men i nästa andetag presenterar han likväl ett antal sådana (s 47-51). Nämnde Stephen Meyer gör i sin bok Signaturen i Cellen den intressanta observationen att de flesta av de kriterier som brukar användas för att förklara ID ovetenskaplig är tveeggade svärd – de kan lika gärna vändas mot evolutionsteorin. Ta som exempel det här med falsifierbarheten (s 47) – att en teori måste gå att motbevisa. Vad skulle kunna motbevisa evolutionen?
E. föreslår att ett påträffat kaninfossil i ett kambriskt berglager skulle göra det. Men i praktiken vet han lika bra som jag att det inte skulle ha någon som helst påverkan på evolutionsteorins trovärdighet. Ett sådant fossilfynd skulle helt enkelt betraktas som en anomali (en avvikelse från det normala) som framtida forskning säkert kommer att kunna förklara. Tills vidare sätter man helt enkelt en parentes runt det och låter evolutionståget tuffa på som vanligt och fortsätter att hävda som E. att
[m]iljoner fossil har grävts upp, identifierats och daterats, men ännu så länge har man inte hittat någonting som klart och tydligt ligger fel. (s 47)
Fel i förhållande till vaddå, kan man undra? E. tycks glömma att det inte är evolutionsteorin som förutsagt den ordning som alla fossil ligger i. Det är den ordning i vilken man hittar fossilen som ligger till grund för den ordning i vilken evolutionsteorin säger att de blev till. När man med jämna mellanrum hittar fossil i ”fel lager”, ja då skrivs den evolutionära historien om. Vi läser om det i media stup i kvarten. Så det där citatet av E. betyder i praktiken ingenting, det är tom retorik.
Jag är inte ensam om att ifrågasätta bristerna i evolutionsteorins falsifierbarhet. Forskare som konstaterat samma sak är Karl Popper (vetenskapsfilosof,1992-1994), Imre Lakatos (vetenskapsfilosof, 1922-1974), Thomas Kuhn (vetenskapshistoriker, 1922-1996), Paul Feyerabend (vetenskapsfilosof, 1924-1994), Stephen Jay Gould (paleontolog, evolutionsteoretiker, 1941-2002) och Richard Lewontin (populationsgenetiker, 1929-2021)
De argumentationskriterier som E. listar (s 49-50) tillhör samma kategori som falsifieringskriteriet, det vill säga det kan lika gärna vändas mot försvarare av evolutionsteorin.
E. tar även upp etableringskriteriet (s 50). Det är att forskare måste ha relevant utbildning, bedriva sin forskning på etablerade universitet eller företag och publicera sina referentgranskade slutsatser i betrodda tidskrifter.
ID-forskares kvalifikationer är inget problem. Däremot är det ett uppmärksammat problem att det är ytterst svårt för forskare som är öppna med att de är positivt inställda till ID att få möjlighet att bedriva sin forskning på statligt finansierade universitet, och om möjligt ännu svårare att få sina resultat referentgranskade på grund av den aktiva censur som tidskrifternas redaktörer bedriver. Den redaktör som råkar släppa igenom en ID-vänlig artikel till referentgranskning och publicering löper stor risk att bli av med jobbet. Det här är ett väl belagt fenomen.
I ärlighetens namn är det inte bara ID-folk och kreationister som får känna av det här. Det är många andra forskare som vittnar om samma sak och som är djupt kritiska till rådande referentgransknings- och publiceringsrutiner. Det är många som under åren fått se sina nya teorier ratade och först långt senare fått upprättelse. Några exempel:
• Kontinentaldriftsteorin (Wegener 1912)
• Transposoner (Barbara McClintock, 1967)
• Endosymbiontteorin (Lynn Margulis, 1967)
• Prioner (Stanley Prusiner, 1980-talet)
• Kvasikristaller (Dan Shechtman, 1982)
Kort sagt: Det är oftast bra med referentgranskning, men proceduren kan också bidra till att konservera rådande teorier och därigenom hämma den vetenskapliga utvecklingen.
Grunden för naturvetenskapen
Naturvetenskapen bygger på ett antal premisser, som till exempel att naturlagarna existerar och att universum är enhetligt, det vill säga att lagarna inte bara gäller här på jorden utan också i avlägsna galaxer. Andra förutsättningar är att vi kan lita på våra sinnen och på våra intellektuella förmågor som logik och matematik, inberäknat metoder som adbuktionsbevis, deduktionsbevis och mycket annat som E. tar upp i den här första avdelningen av boken. Allt sådant där tar han för givet.
Men varför skulle det vara självklart utifrån en naturalistisk syn på världen där allting har skapat sig själv utan plan, mål och mening? Ska man vara krass så kan inte den naturalistiska världsbilden motivera varför universum skulle vara rationellt och begripligt över huvud taget.
Och samtidigt är allt sådant en nödvändig rekvisita för vetenskaplig verksamhet. E. ”lånar” den från den bibliska världsbilden utan att be om lov. Bildligt talat spelar han kort, vetenskapsspelet, men struntar i att fråga sig varifrån kortleken kom. Den är en del av verkligheten som han inte tycker behöver förklaras.
Men naturligtvis behöver en trovärdig världsbild kunna förklara även sådana saker. En sådan förklaring hittar vi i den bibliska världsbilden. Och endast där! Läs mer i Appendix 4.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 8. Evolutionsteorin
Efter de vetenskapsteoretiska kapitlen följer ett kapitel om evolutionsteorin. Det är naturligtvis viktigt för E. att ha ett sådant kapitel och argumentera för evolutionsteorins trovärdighet. Varför då? Jo, utan det hade hans bok i stort sett bara innehållit kritik mot ett annat synsätt än hans eget och det är ju just det som han klandrar företrädare för ID och kreationism för att göra: att vi bara kritiserar evolutionsteorin och inte har någon egen positiv evidens att visa upp för vår hållning. Det här är en allvarlig anklagelse, så håll i dig – det här kommer att bli dokumentets längsta kapitel.
Centralt för E., liksom för de flesta evolutionsbiologer, är att evolutionen är planlös, den är ateleologisk, vilket innebär att den saknar syfte och mål. Redan i början av evolutionskapitlet betonar han därför att evolutionen
inte strävar mot något fjärran mål, som att gå ’framåt’ mot alltmer komplexa organismer. (s 52)
Han nämner parasiter som exempel på motsatsen.10 (s 52) Mer komplex eller mindre komplex, framåt eller bakåt, bra eller dåligt – evolutionsteorin är helt oberoende av sådana mänskliga värderingar – evolutionen helt enkelt bara ”är”.
Här är det på sin plats att nämna att det ovannämnda resonemanget är en halvsanning, eller kanske snarare en sanning som E. använder i en medveten eller omedveten strategi för att förflytta diskussionens fokus från det centrala till det perifera.
Evolutionen är nämligen först och främst en modell som gör anspråk på att förklara hur en ensam liten individ av en primitiv mikroorganism, LUCA (Last Universal Common Ancestor), under en period av så där fyra miljarder år har genomgått förändringar som förvandlat dess ättlingar till alla de miljontals livsformer, inklusive oss själva, som jorden vimlar av idag och som befolkat den under tidigare tidsepoker. Detta är den centrala fråga som evolutionsteorin behöver besvara på ett trovärdigt sätt. Alltså hur har den genetiska information som kodar för alla livets strukturer, organ och egenskaper utvecklats? Inte hur den nu och då kan avvecklas genom parasitism.
Det här är ett smart drag av E. Den vetenskapliga litteraturen är nämligen överfull av evidens för att organismer regelmässigt avvecklar diverse egenskaper som är överflödiga för dem i vissa sammanhang. Genom att definiera sådant som evolution (utveckling) i stället för att kalla det vid dess rätta namn, devolution (avveckling) behöver han inte bekymra sig om att hitta konkreta exempel på att organismer förvärvar kvalitativt nya egenskaper och strukturer. Han kan i stället peka på exemplen på avveckling som evidens för att evolutionen är ett faktum. Svart blir vitt.
Att känna till hur E. tänker här är en nyckel till att förstå hans sätt att resonera när han sedan behandlar evolutionsmekanismerna. Låt oss nu koncentrera oss på dem en stund.
Variation
Variation är en av dem. E. har det som en egen underrubrik (s 54). Hans syfte är att förklara för läsaren hur nya genvarianter uppstår i populationer av levande organismer. Han nämner de båda mekanismerna mutationer och rekombination. Det förstnämnda är när det sker slumpmässiga förändringar i en sekvens av nukleotider (”bokstäver”) i en organisms DNA. Det andra är när genvarianter byter plats med varandra på homologa11 kromosomer i samband med att det bildas ägg och spermier.
E.s budskap är att det är genom de här båda mekanismerna som genetisk variation ökar i en population. Ibland, tillägger han (s 54), kan en kromosom gå av så att de båda delarna av en viss gen sedan bidrar till att öka variationen ytterligare. Det stämmer, men som vi snart ska se med reservation (punkt 2 nedan).
Vidare konstaterar han mycket riktigt att många mutationer är skadliga och att bara ett fåtal är klart gynnsamma. Sedan följer ett uttalande som är värt att analysera lite närmare:
Flertalet mutationer är ’nära-neutrala’, vilket betyder att de bara har små, om ens några effekter. Emellertid kommer de att öka den genetiska variationen i populationen. (s 55)
Det intressanta är inte så mycket vad E. säger, utan hur fakta presenteras och vad kontentan blir för en oinsatt läsare. Den blir följande (rätta mig gärna om du tycker att jag har fel):
Ibland är mutationer klart fördelaktiga. Visserligen är de ofta skadliga, men det gör inte så mycket i praktiken eftersom det stora flertalet mutationer knappast har någon effekt alls utan har den positiva konsekvensen att de ökar populationens genetiska variation.
Jag vill understryka att detta inte är ett citat av E. Men jag är tämligen säker på att han skulle hålla med om min tolkning.
Jag måste säga att jag är imponerad av E.s retoriska skicklighet. Han får här att ett av evolutionsteorins allvarligaste problem att framstå som en tillgång utan att uttala en enda direkt osanning! Låt mig förklara.
E. använder två strategier. Den första är att han i flera fall töjer på sanningen och den andra att han undanhåller läsaren viktiga fakta:
1. Fördelaktiga mutationers sällsynthet
E. skriver helt korrekt att det är få mutationer som är fördelaktiga (s 55). Men han skriver ingenting om hur ovanliga de är.
Den bäst studerade organismen är den vanliga tarmbakterien Escherichia coli (E. coli). Den kan under gynnsamma förhållanden bilda en ny generation på 20 minuter. Man har följt en och samma bakteriepopulations genetik under 80 000 generationer. Andelen fördelaktiga mutationer uppskattas till någonstans i intervallet en per 105 till 106 eller mer, alltså ungefär en på miljonen!13
Dessa uppskattningar gäller bakterier som aktivt använder sig av mutationer som en aktiv överlevnadsstrategi, det vill säga under cellens kontroll. Jag har tidigare skrivit om detta i magasinet Genesis.13 När det gäller till exempel människor med en helt annan komplexitetsnivå på arvsmassan finns det skäl att förmoda att andelen är avsevärt lägre än en på miljonen.14
Dessutom bör det tilläggas att den absoluta merparten av mutationer som innebär en fördel för sin bärare är degenerativa, det vill säga nedbrytande till sin karaktär. Det formulerades träffande av mikrobiologen Marcia Stone i hennes artikel i tidskriften Bioscience: 15
alltså: ”för mikroorganismer är avveckling lika med utveckling! Richard Lenskis världsberömda långtidsförsök med E. coli har tydliggjort bakteriernas strategi. Den kan jämföras med en person som är ute och kör bil och till sin fasa upptäcker att bensinen håller på att ta slut. Han är tio mil från närmaste bensinstation (Norrland) och bensinmätaren visar att han bara har bensin för nio mil. Vad göra? Han bestämmer sig för en drastisk åtgärd. Han stannar bilen och lastar av allt bagage tillsammans med reservhjulet. Sedan demonterar han baksätet och passagerarsätet och ställer dem i diket. Sedan fortsätter han sin färd, och med den nu betydligt lättare bilen når han fram till bensinstationen och kan tanka upp sin bil. Sedan kör han tillbaka och hämtar upp sina pinaler och ordningen är återställd. Det har visat sig att Lenskis bakterier gör precis så i en stressituation – de gör sig av med genetiskt material som inte är nödvändigt för att överleva.16 När miljön återgår till det normala kan de importera de förlorade generna från en annan bakterieindivid.17 Hur är det med oss människor då? Det klassiska exemplet från skolornas läroböcker med den fördelaktiga mutationen i form av anlaget för sicklecell-anemi som ger sin bärare en viss immunitet mot malaria illustrerar samma fenomen: det är onekligen en fördel med en muterad gen som gjorde det möjligt att överleva spädbarnsåren tack vare att man slapp drabbas av malaria. Men det är på samma gång en tydlig nackdel att bära på en ärftlig blodbristsjukdom vilket är den andra sidan av myntet. En fördelaktig mutation under vissa omständigheter (när man bor i ett malariadrabbat område), men till skada under alla andra omständigheter. Jag vet sedan tidigare dialoger18 med E. att han har starka invändningar mot att jag gör skillnad mellan att en mutation är fördelaktig och att den är konstruktiv, uppbyggande, till sin karaktär. E. tycker inte att det spelar någon som helst roll, eftersom han ser evolutionen som ”blind” för våra mänskliga värderingar. Du som läser det här får själv avgöra vem av oss som resonerar klokast. Jag påstår att eftersom evolutionen framför allt ska förklara utvecklingen från en primitiv mikroorganism till en människa19 och eftersom en människa har oräkneligt många egenskaper som denna mikroorganism saknade, så måste en sådan utveckling ha varit en ytterst konstruktiv process bestående av oräkneliga mängder av just konstruktiva förändringar. Det räcker med andra ord inte att förändringarna (mutationerna) är fördelaktiga i samma bemärkelse som exemplen med bilen och sickle-cell-anemin ovan illustrerar. Man kan inte göra sig en förmögenhet på att öka försäljningen om man går en krona back på varje försåld enhet. Men E. tänker inte så. Eftersom han tycker att det är lika mycket evolution när en organism förlorar gener och organ som när de (hypotetiskt) tillkommer, och eftersom han inte vill kännas vid skillnaden mellan fördelaktiga och konstruktiva mutationer så är det för honom ett pseudoresonemang. 2. Överkorsning inuti gener är skadliga E. menar (s 54) att en gen som delas genom överkorsning (rekombination) bidrar till den genetiska variationen. Det är sant, men det finns inga välbelagda exempel där partiella genvarianter skapade direkt av överkorsning mitt i en gen tydligt och markant har visats öka fitness i naturliga populationer. Däremot orsakar de en rad allvarliga sjukdomstillstånd, däribland blödarsjuka. 3. Mutationer är överlag skadliga E. nämner att många mutationer är skadliga, men han kommenterar det inte. Det är viktigare för honom att sätta strålkastarbelysningen på de nära-neutrala mutationerna, som vi snart kommer till. Men här måste vi vrida om strålkastarna från spekulationernas område till den krassa verkligheten. Vilken verklighet talar jag om nu? Jo, det faktum som vi alla känner till: mutationer är bokstavligen talat livsfarliga! Din tandläkare vet om det. Det är just därför som han gömmer sig bakom en vägg innan han trycker på röntgenknappen. Han vill inte utveckla cancer eller få förändrade spermier på grund av upprepade strålningsdoser så att hans barn och barnbarn får någon av de miljontals genetiska defekter som mutationer bevisligen förorsakar. Voffo e de på detta vise? frågade rumpnissarna i Ronja Rövardotter. Svaret är enkelt. Vi vet att sannolikheten för att slumpmässiga förändringar i ett informationssystem (vilket DNA är) skadar information är oerhört mycket större än sannolikheten för att de åstadkommer någonting bra, och i synnerhet någonting konstruktivt. Detta är anledningen till den skeva fördelningen mellan positiva/konstruktiva och negativa/nedbrytande mutationer som vi nyss noterade råder i den levande världen. Det är en tydlig indikation på att vår arvsmassa inte till stora delar är ett evolutionärt skrotupplag (begreppet ”skräp-DNA” kommer jag att behandla senare, eftersom det är ett ID-argument som E. vill avfärda) utan just ett informationssystem. Hela diskussionen med mutationers skadlighet har givetvis stor bäring på om vår arvsmassa är skräp eller inte. Att röra om i en hög med skräp är ju ofarligt, men gör man det i operativsystemet på en dator så är läget förstås ett annat. Men, som sagt, E. finner ingen anledning att kommentera det allting överskuggande faktum att mutationer är den mest destruktiva av alla naturliga processer (utom som sagt om man råkar vara en bacill eller B-lymfocyt, vilket få av oss är). E. förutsätter att en stor andel av de mutationer som en organism har ingen eller bara obetydliga effekter. Men det kan vara väldigt svårt att veta. En mutation som inte har någon speciell inverkan i en viss celltyp kan ha en dramatisk inverkan i en annan icke-studerad celltyp. Eller en lika dramatisk effekt i ett visst skede i embryonalutvecklingen eller under en speciell typ av miljöförhållanden. Så det enda vi kan säga med någorlunda säkerhet är att skadeeffekterna är större än hittills blivit dokumenterat. 4. Nästan-neutral är inte nyttig Låt oss nu vrida tillbaka strålkastaren dit E. hade ställt den från början – mot de nästan-neutrala mutationerna. I dem ser han sin räddare i nöden. De kommer ju med hans ordval att For microbes, Devolution is Evolution
öka den genetiska variationen i populationen. (s 55)
Tankegången här är att organismerna över tid samlar på sig mängder av muterade gener med
små, om ens några effekter (s 55)
som sedan kan användas som ett råmaterial för det naturliga urvalet att verka på. De ”små effekterna” kan förstärkas och utvecklas till allehanda viktiga funktioner E. framställer alltså nästan-neutrala, svagt bra och svagt dåliga, mutationer som en positiv potential för framtida förändring. Är det verkligen så?
Det är verkligen inte så! Det första vi bör uppmärksamma är fördelningen mellan de svagt skadliga och de svagt fördelaktiga mutationerna. Av E.s sätt att uttrycka saken får man intrycket att det skulle vara någon sorts 50/50-fördelning av dem. Men vad finns det för anledning att förvänta sig att vi skulle ha en helt annorlunda fördelning bland mutationerna med svaga effekter än fördelningen bland de med mer påtagliga effekter? Naturligtvis ingen alls – andelen svagt fördelaktiga mutationer bör rimligen ligga någonstans kring en på miljonen även här.
Det är alltså ingen neutral massa av mutationer utan stora mängder av till övervägande delen svagt negativa sådana som ansamlas i organismernas arvsmassa just därför att det naturliga urvalet inte ”kommer åt” att eliminera de individer som är bärare av dem. Genetikern John C. Sanford har skrivit en mycket intressant bok med titeln Genetic Entropy20 där han utvecklar just detta förhållande. Det faktum att varje ny generation av människor har ungefär 100 nya mutationer i förhållande till sina föräldrar (vilket E. mycket riktigt konstaterar (s 55), parat med faktumet att det går minst en miljon svagt skadliga på varje svagt fördelaktig innebär i praktiken att varje ny generation av människor (och inget säger att situationen skulle vara stort annorlunda hos andra organismer) ackumulerar hundra mer eller mindre skadliga mutationer. Det här har med nödvändighet en nedbrytande effekt på organismers arvsmassa över tid. Sanford kallar det ökande genetisk entropi (oordning). I Appendix 1 förklarar jag vad det beror på att det naturliga urvalet inte klarar av att hantera de nära-neutrala mutationerna.
Evolutionsgenetikern Alexey Kondrashov sammanfattade problematiken i rubriken på sin artikel i Journal of Theoretical Biology:
Contamination of the genome by very slightly deleterious mutations: why have we not died 100 times over?” (Kontaminering av arvsmassan genom mycket svagt skadliga mutationer: varför har vi inte dött hundra gånger om?).21
Innan vi lämnar avsnittet med variationsskapande mekanismer så kan vi ställa oss frågan: Om det nu är så uppenbart som jag hävdar att evidensen visar, att mutationer är övervägande nedbrytande till sin karaktär – hur kan då E. låta påskina motsatsen?
Det handlar inte om – eller jag vill absolut inte tro – att E. medvetet försöker undanhålla läsaren viktiga fakta. Det är bara det att hans totala övertygelse om evolutionens faktum gör att han medvetet eller omedvetet avfärdar vissa delar av verkligheten, likt de ovannämnda, som mindre intressanta och mindre relevanta. Jag tror att han tänker så här: Vi människor existerar ju bevisligen, liksom alla andra jordens varelser. Följaktligen måste mutationer vara nyskapande och konstruktiva (även om han aldrig skulle använda sig av det ordvalet). Och eftersom evolutionens råmaterial omöjligen kan vara de starkt skadliga mutationerna och inte heller bara de tydligt fördelaktiga eftersom de är så få till antalet, så måste hemligheten ligga hos de nära-neutrala mutationerna. Att merparten av dem är svagt negativa bekymrar honom inte, för om miljön förändras så är det teoretiskt möjligt att några av dem i stället blir svagt fördelaktiga.
Det här resonemanget utgår förstås inte från någon empirisk evidens, utan är ett spekulativt och bakvänt resonemang med evolutionens “faktum” som utgångspunkt.
5. Rekombination är inte slumpmässig
Rekombination är långtifrån den slumpmässiga process E. vill få det att framstå som. Vi vet idag att det handlar om en ytterst komplex och noggrant reglerad process, annars skulle en cell aldrig överleva en celldelning.
Robusthet
Under nästa rubrik (s 55) vill E. förklara för oss hur viktigt det är att muterade gener inte avlägsnas från populationen av det naturliga urvalet, för då skulle det inte kunna bli någon evolution. Det här beror, menar han, på samma mekanism som gör att vi kan vara bärare av skadliga genvarianter utan att påverkas så mycket av dem. Den mekanismen är cellers och organismers robusthet (s 55). Det finns bildligt talat hängslen och livremmar (så kallad redundans) i livets kemi som dämpar mutationernas skadeverkningar.
Det här stämmer, men återigen har vi en tolkning utifrån E.s evolutionära grundsyn. Livets olika system är utan tvivel robusta, men finns det fler sätt att betrakta dem på?
Ja, det gör det. E. anser att robusthet är en egenskap hos levande organismer som utvecklats därför att den medger evolverbarhet, möjlighet att utvecklas. Det där tangerar förstås ett cirkelresonemang, men låt oss bortse från det och helt enkelt se det som en tänkbar förklaring.
Designperspektivet på livets robusthet
Det finns ett alternativt sätt att tolka naturens redundans och det är utifrån ett designperspektiv. I vår bil finns det dubbla bromskretsar som gör att om något problem skulle slå ut den ena kretsen, så kommer bilen ändå att bromsa in när vi trycker ned bromspedalen. Bilen är alltså också utrustad med både hängslen och livrem. I flygplan finns det rentav tre separata styrsystem, därför att konsekvenserna skulle kunna bli ännu allvarligare i det fallet. Det här med redundans är alltså en designprincip som ingenjörer tillämpar för att deras konstruktioner ska vara så pålitliga och säkra som möjligt.22
Tack vare samma princip i våra kroppar kan vi punktera ena lungan och ändå överleva och bli återställda, om en njure råkar bli sönderslagen kan den friska öka sin kapacitet och ta över den trasigas funktion, och så vidare.
Enligt designperspektivet använde sig vår skapare av den här principen med inbyggd redundans långt innan våra ingenjörer hade kommit på tanken. Det är inte alls självklart att evolutionen skulle vara lika förutseende - den är ju faktiskt både blind och planlös, eller hur?
E.s slutsats är att robustheten i kombination med de nära-neutrala mutationerna leder till att de senare ansamlas i de levande organismerna över tid. Det är helt sant. E. ser det som en nödvändig förutsättning som möjliggör evolutionen. Designförespråkare ser det som en destruktiv process som långsamt men säkert förstör levande varelser. Kontrasten i synsätt kan inte vara större. Vem har rätt? Jag är säker på att även en lekman kan avgöra det genom att ställa den grundläggande frågan: Är mutationer någonting bra eller någonting dåligt? Låt evidensen tala för sig själv. Ge akt på din tandläkare.
Selektion
Att mutationer ständigt skapar nytt råmaterial för evolutionen i form av muterade gener är själva fundamentet för evolutionsteorin. Ytterst finns det ingen annan mekanism för variation. Visst kan anlag (genvarianter) omfördelas och uppträda i nya kombinationer genom olika slag av rekombination,23 men innan man kan blanda, kupera och dela ut sina spelkort måste man ha en kortlek på bordet. Det är mutationerna som anses ha skapat den.
Vi såg nyss (punkterna 1-5 ovan) att det inte är självklart att mutationer är så kreativa som E. försöker få dem att verka, men låt oss för en stund bortse från det. Låt oss i stället analysera vad han skriver om nästa grundpelare inom evolutionsteorin, nämligen det naturliga urvalet, selektionen, (s 56) det som var och fortfarande är den bärande idén i Charles Darwins utvecklingslära.
Selektion – principen att de bäst anpassade individerna i en population har bäst chanser att föra sina gener vidare till nästa generation – är som idé helt genial. Den kan förklara precis allting, speciellt när den tillämpas i form av det närliggande begreppet selektionstryck. E. berättar hur de olika formerna av selektionstryck kan få en population att utvecklas i en viss riktning – eller ingen alls – beroende på miljöomständigheterna.
När vi ser naturfilmer hör vi nästan alltid programledaren lovorda det naturliga urvalet som under årmiljonerna slipat fram allt vackert och finurligt i naturen. Det är då klokt att påminna sig själv om att urvalet i praktiken inte kan skapa någonting alls på egen hand – det kan bara göra en enda sak: eliminera de individer som är mindre lyckligt lottade, och eventuellt på längre sikt, de misslyckade gener som de är bärare av. Sett ur det perspektivet förstår vi att det är mutationerna och inte samspelet med miljön som utgör evolutionens främsta flaskhals. Men det är främmande för evolutionsbiologer att tänka på det viset.
Det finns särskilt en formulering i E.s redogörelse för selektion som är viktig att belysa. På s 57 skriver han:
Selektion medför därför en minskad genetisk variation i populationen. Emellertid kommer mutationer att leda till att den genetiska variationen åter ökar.
Den första meningen är ett helt fundamentalt och väldokumenterat förhållande, som i sig innehåller en oerhört stor utmaning för evolutionsteorin: Selektionen, den mekanism som anses vara evolutionsteorins främsta drivkraft, leder till att den genetiska variationen – eller om man så vill den biologiska mångfalden inom en population – minskar i stället för ökar! Varför är det så?
Det handlar om grundläggande mendelsk genetik. Alla former av riktad selektion handlar om att vissa befintliga alleler (genvarianter) blir vanligare i en population medan andra blir ovanligare. Efter ett antal generationer kommer vissa alleler att fixeras i populationen så att alla individer bär dem. Om det låter kryptiskt så läs artikeln i not 24.
Det går att illustrera det här förhållandet utan att behöva blanda in några avancerade teoretiska resonemang. Tänk på våra hundar. Det finns omkring 500 olika hundraser i världen (och naturligtvis ofantliga mängder av hybrider dem emellan). De har alla vargen som stamfader. Människan har ”format” dessa genom genomtänkt selektion. Man har helt enkelt valt ut valpar med de egenskaper man föredragit för att få fram hundvarianter som lämpar sig för det ändamål man önskar – jakthundar, vallhundar eller knähundar. Under den här urvalsprocessen har man sett till att individer med mindre önskade egenskaper inte har fått möjlighet att föröka sig. Man har alltså från ett djur med generella egenskaper (vargen) åstadkommit en specialisering i form av en stor mängd specialiserade djur.
Det här förhållandet var faktiskt någonting som var en central del av Charles Darwins resonemang i hans berömda bok Om arternas uppkomst. Om människor på bara några hundra år kunnat selektera fram alla möjliga slags husdjur – tänk då på vad naturen själv kan åstadkomma under miljontals år.
Det finns förstås en liten hake i Darwins resonemang. Det är nämligen inte på något vis självklart att ointelligenta, viljelösa naturliga processer äger samma förmåga som intelligenta människor att forma ett levande material.
Men alldeles oavsett om det är människan eller ”naturen själv” som står för selektionen och specialiseringen, så är det likväl så att konsekvensen är minskad genetisk mångfald. I fallet med hundar kan vem som helst dra den slutsatsen utifrån två helt uppenbara observationer:
För det första kan en pudel inte överleva ens en svensk vinter i det fria. Varför inte? Ju under selektionen har mängder av genvarianter som en gång fanns i den ursprungliga vargen eliminerats längs vägen. Biologiskt kan det formulerats som att tamhundarna förlorat i heterozygoti och blivit mer homozygota. De genvarianter som gav och ger vargen dess luktsinne, jaktegenskaper, uthållighet och allmänna tålighet finns inte längre kvar i pudelgenomet. Selektion utarmar.
För det andra: Ett par pudlar kommer generation efter generation att bara generera nya kullar av – pudlar. Vargens alla genvarianter som utgör potentialen till att bilda hundratals specialiserade djur med vitt skilda egenskaper har gått förlorad längs vägen. Återigen: selektion utarmar.
När jag läser E.s formulering får jag känslan att han inser att det motsägelsefulla i att selektionen gör populationer genetiskt fattigare, och därför bestämmer sig för att lägga till en mening för att rätta upp situationen, nämligen den om att mutationerna kommer att öka variationen på nytt.
Men finns det verkligen evidens som stöder den tanken? Nej, tvärtom. Världens snabbaste landdjur illustrerar det. Den nutida populationen av geparder (Acinonyx jubatus) har en extremt liten genetisk variation, de är praktiskt taget ”enäggsyskon”. Det visar att de någon gång under historien genomgått en så kallad genetisk flaskhals då bara några få individer, kanske ett ensamt par, överlevde. Från dessa har det sedan dess bildats en ny population. Om E. haft rätt borde populationen under de senaste årtusendena ha samlat på sig betydande mängder av nya gener som skulle ha skapat en ny variation inom arten. Men så är uppenbarligen inte fallet. E. har fel. Mutationer kan inte kompensera för selektionens utarmande effekt.
Mutationer har förstås en påverkan, men det sätt på vilket de påverkar organismerna illustreras också tydligt i hundarnas (artificiella) utveckling. Den korta nosen hos en boxer eller bulldogg är resultat av sådana. Mutationer med den sedvanliga konsekvensen av försämrade egenskaper. De har inte bara gett hundarna sämre luktsinne25 utan även andningssvårigheter.
E.s sätt att beskriva selektionen är verkligen att töja på sanningen. Ja, mutationer tillför ny variation, men dels är denna variation praktiskt taget alltid till skada för organismerna, och dels kan de inte uppväga selektionens utarmande verkan på arvsmassan. Hur kan jag vara så säker på det? Svaret är elementärt: Annars hade ju inte den väldokumenterade observationen att selektion minskar den genetiska variationen kunnat göras i första rummet.
Selektionsresonemanget avrundas med ett exempel (s 57) på hur en skadlig mutation (M) kan elimineras genom rekombination och innebära en fördel för en individ nästa generation. Jag förstår varför han har med exemplet, för någonstans inser han att det här med mutationernas skadlighet är ett dilemma för evolutionsteorin. Det här är ett sätt att mildra problemet
Men hur realistiskt är det mot bakgrund av vad vi vet om fördelningen av fördelaktiga och skadliga mutationer? Notera att i E.s exempel finns en kromosom med tre anlag, W, M och Br. Det illustreras som i figuren (överst på s 55):
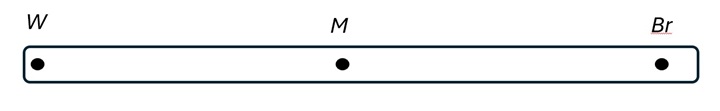
M är som sagt en skadlig genvariant. Men W och Br är båda gynnsamma genvarianter. E. målar alltså upp ett scenario där ett skadligt anlag har råkat hamna i samma kromosom som två gynnsamma anlag. Du minns kanske att andelen fördelaktiga mutationer är runtn en på miljonen. En situation som i E.s scenario har alltså en sannolikhet på ungefär en på 1012.
Hittills kan vi alltså konstatera att E. erkänner att mutationer oftare är skadliga än fördelaktiga och att det naturliga urvalet gör populationer genetiskt fattigare. Samt att han misslyckas med sina försök att bortförklara de fundamentala problem detta innebär för evolutionsteorin.
Jag vill poängtera att det ovan sagda inte får missförstås som att det naturliga urvalet inte skulle ha några positiva effekter alls i naturen. I nuvarande26 ekosystem har exempelvis förekomsten av rovdjur en konserverande effekt på populationers livskraft genom att svagare individer elimineras i högre grad än friska. Gäddans huggreflex utlöses till exempel av ett fiskedrag som imiterar en skadad bytesfisk. Selektion är med andra ord en mekanism som kan dämpa det genetiska sönderfallet (entropiökningen) på grund av mutationerna, men inte bidra till någon kvalitativ utveckling. Selektion kan bidra till att finkalibrera organismerna till deras miljö, men sannolikt i mindre omfattning än evolutionsbiologer menar. Så kallade epigenetiska faktorer är sannolikt viktigare i sammanhanget (se rubriken Epigenetik strax nedan).
Punkterad jämvikt
Nästa rubrik är Punkterad jämvikt. (s 57) Det är en teori som formulerades i början av 1970-talet av de båda paleontologerna (fossilforskarna) Stephen Jay Gould och Niles Eldredge. Avsikten med teorin var att förklara hur det kommer sig att man inte återfinner stegvisa övergångar mellan livsformerna i fossilarkiven. I stället kännetecknas fossilen av plötsliga uppträdanden följda av det man tolkar som långa perioder av stasis när organismerna inte tycks förändras nämnvärt. Ett numera klassiskt citat av Gould från 1977 är:
Den extrema sällsyntheten av övergångsformer i fossilens arkiv är och förblir paleontologins affärshemlighet. 27
Teorin går ut på att populationer kan leva långa tider under stabiliserande selektion. Om sedan miljön förändras snabbt, speciellt i en liten delpopulation, kan det uppstå ett plötsligt selektionstryck med riktad selektion så att det sker förhållandevis mycket snabba förändringar inom den lilla populationen som senare sprids i den större. Tack vare att populationen där de förhållandevis snabba förändringarna sker är liten och geografiskt begränsad är sannolikheten liten för att det bildas några fossil som dokumenterar förändringen, och det uppstår en skenbar lucka i fossilserien av organismen i fråga. E. konstaterar:
Man kan få höra att punkterad jämvikt skulle utgöra ett problem för evolutionsteorin, men det stämmer inte – fenomenet ryms helt inom teorin. (s 58)
Men jag har varit med under alla de år som gått sedan teorin presenterades, och jag har aldrig någonsin hört argumentet att teorin skulle vara ett problem för evolutionsteorin. Det där måste E. ha fått om bakfoten.
Vad vi ID-anhängare och kreationister däremot har hävdat, och fortfarande hävdar, är att teorin inte besvarar frågan varför mellanformer systematiskt saknas i fossilen. Det främsta skälet till det är att selektion, och i synnerhet snabb sådan i en liten population, leder till en snabb och ofta drastisk minskning av den genetiska diversiteten.28 Detta är ett väldokumenterat fenomen som kallas genetisk drift. Det kan visserligen leda till snabba förändringar av organismerna, men den genetiska bakgrunden är avveckling och inte utveckling. Som tidigare påpekats är det det senare som evolutionsteorin behöver förklara, inte det förra. Teorin om punkterad jämvikt är alltså ingen räddare i nöden. Faktiskt precis tvärt om.
Neutral evolution
Neutral evolution (s 58) är teorin att mycket av den variation som finns i naturen inte är ett resultat av mutationer med efterföljande selektion, utan mutationer utan selektion. E. har en underrubrik om saken och skriver:
Det betyder att inte alla egenskaper hos en organism måste finnas där för att de har en adaptiv funktion – de kan bara ”råka” finnas där… (s 58)
Ibland, menar E., skapar mutationer alltså variation utan att det är någon selektion som verkar på dem, och nämner bladformerna i Michael Dentons film Biology of the Baroque29
Vi har vid det här laget konstaterat hur problematiskt det att förklara uppkomsten av biologiska strukturer och funktioner genom en kombination av mutationer och naturligt urval. Skulle det vara lättare att förklara uppkomsten av dem utan någon selektion? Då finns det ju ingen trend på förändringarna i någon särskild riktning. Idén bygger alltså på att enbart slumpmässiga mutationer förmår bygga nya gener för nya egenskaper i frånvaro av selektion. Finns det evidens för någonting sådant? Ingen alls. Välkommen att bevisa motsatsen.
Epigenetik
E. tar sedan upp ett tämligen nytt område inom biologin, nämligen epigenetiken. (s 58) Det är faktorer som styr embryots utveckling och kan leda till snabb artbildning utan att ordningen på DNA-bokstäverna ändras. Det är alltså en dimension ”ovanpå” (epi-) DNA som styr och reglar den på sätt som idag är mycket ofullständigt utforskade. Anledningen till att han tar upp ämnet är att ID-förespråkare och kreationister menar att den nya vetenskapsgrenen utgör ett problem för evolutionsteorin. Men det tycker inte E.;
Detta är inte riktigt. Den naturliga selektionen verkar på alla egenskaper, oberoende av om de skulle kontrolleras av genetiska eller epigenetiska mekanismer. (s 59)
Men det finns ett förhållande som jag misstänker att E. inte känner till.30 Det är att en av de bäst kända epigenetiska mekanismerna31 leder till att DNA-molekylen åldras. Åldrandet går till så, att kvävebasen C (cytosin) omvandlas till T (tymin). Konsekvensen är att arvsmassan för varje ny generation förlorar G-C par till förmån för A-T-par. Och processen är delvis irreversibel, det vill säga omöjlig för cellen att reparera.32 Det här leder till att komplexiteten i organismers arvsmassa minskar sakta men säkert för varje ny generation, och det späder på den genetiska entropiökningen som jag tidigare beskrivit.33
Låt mig påminna om att evolutionsteorin behöver kunna förklara hur de levande organismernas arvsmassa, alltså de sekvenser i DNA som alla organismer har, har uppkommit. Vi har redan sett hur mutationer är en ytterst tveksam källa till variation för komplexa organismer. Vi har också sett hur det naturliga urvalet saknar tillräcklig ”synskärpa” för att komma åt det stora flertalet av mutationer som därmed ansamlas i arvsmassan. Epigenetiken bidrar till att minska synskärpan ytterligare genom att en stor mängd av variationen i naturen, och även artbildning,34 beror på faktorer som inte har med ordningsföljden av DNA-bokstäverna att göra.
Det här motsäger E.s påstående att epigenetiken inte utgör något problem för evolutionen.
Inget nytt under solen
I en debatt med E. för en del år sedan35 påminde jag honom om den holländske botanikern Hugo de Vries som på sin tid sade att naturligt urval är utmärkt på att förklara ”survival of the fittest”, men däremot inte ”arrival of the fittest”. Alltså att selektionen är bra på att anpassa organismer och deras egenskaper till sin aktuella miljö men dålig på att förklara deras uppkomst, vilket ju är vad teorin gör anspråk på att göra.
Kanske kom E. ihåg det där citatet och förärade det en egen rubrik, eller så fick han det någon annanstans ifrån. Hur som helst, under den här rubriken finns en av bokens mest avslöjande formuleringar som illustrerar hur viktig reduktionismen är i E.s världsuppfattning. Som jag nämnde tidigare så antyder han att han inte är främmande för den filosofiska idén att den komplexitet vi ser i naturen kan reduceras till mer grundläggande, underliggande faktorer. Det kommer till uttryck på några ställen i boken, och här är andra gången.
Bakgrunden är att han citerar Charles Darwin när denne uttrycker sin tro att alla organ i alla organismer kan förklaras genom ett stort antal små förändringar. Det som med ett modernt ord brukar kallas gradualism. E. håller med. Han skriver, snudd på känslosamt:
Och det gör ärligt talat inte jag heller. Inte ett enda fall där något helt nytt dyker upp från ingenstans, något så radikalt nytt att man måste utesluta kända evolutionära mekanismer. Däremot finns det fall där man kan tänka sig att nya funktioner skulle kunna evolveras ur strukturer som redan existerar.” (s 60)
E. syftar på det populära evolutionära begreppet exaptation.36 Det är när en viss egenskap förändras och övergår i en annan egenskap, via ett mellanskede där det eventuellt finns visst mått av båda egenskaperna på samma gång.
Varför är exaptation så populärt bland evolutionsbiologer då? Jo, det beror på att det tycks lösa upp ännu ett gigantiskt evolutionärt dilemma som genom ett slag med trollspöet. E. formulerar dilemmat kristallklart. Jag repeterar. Han känner inte till…
…ett enda fall där något helt nytt dyker upp från ingenstans,
Bästa läsare. Ta nu ett djupt och eftertänksamt andetag och begrunda vad det är denna docent i biologi skriver och dessutom betonar med eftertryck: Han kan, efter ett lång arbetsliv inom de biologiska vetenskaperna, inte påminna sig själv om ett enda exempel på att någonting nytt uppstår i den levande världen!
Docent Lars Johan Erkell hävdar alltså i princip att det inte finns någon kvalitativ skillnad mellan urmikroben och honom själv. Det är helt och hållet en fråga om ”mer av samma”. Det finns en kontinuerlig obruten utvecklingslinje från den första urorganismen till en människa där ingenting helt nytt har dykt upp längs med vägen.
Man kan undra om E. har funderat över om det möjligen kan finnas någon annan orsak än denna till att det verkar vara så helt omöjligt att påvisa tillkomsten av nya strukturer i den biologiska värden? Skulle möjligen design kunna vara ett sådant? Att det finns en Gud som skapade de olika livsformerna i begynnelsen. Nej visst, det är ju ovetenskapligt. Men hur vetenskapligt är då E.:s scenario?
En vanlig läsare av boken bör rimligen dra slutsatsen att om en så kompetent författare som E. uttrycker en sådan här uppfattning så måste det naturligtvis finnas mycket empirisk evidens som bekräftar den. Är det så?
Just detta är den primära frågan: Vad finns det för evidens som pekar i den riktningen? E. har redan tangerat svaret. Han skrev att det finns
fall där man kan tänka sig att nya funktioner skulle kunna evolveras ur strukturer som redan existerar (s 60)
Dels handlar det om evolutionära berättelser med obefintligt eller ytterst omdiskuterad fossil evidens, som till exempel om hur fiskfenor utvecklades till grodben, kräldjursfjäll till fågelfjädrar och så vidare.
Dels handlar det om molekylärbiologiska exempel med enzymmolekyler som får nya funktioner. Vi ska återkomma till det längre fram. E. tar upp ett annat sådant exempel (s 60) där han berättar om hur synförmågan kan ha uppstått. Han menar att det bara krävs två mutationer för att förändra en viss proteinmolekyl så att den kan binda till en annan molekyl än den brukade göra. Nu binder den i stället till en ljuskänslig molekyl (retinal) och vips, så kan den primitiva varelsen se. Sedan behövs det enligt E. inte mycket för att
evolutionen av en alltmer avancerad synförmåga [kan] ta sin början. (s 60)
Han kallar detta
ett fullt rimligt scenario. (s 60)
Hur uppstod synförmågan?
E. vill övertyga läsaren om att allt som behövs för uppkomsten av synförmåga är två slumpmässiga förändringar i DNA-molekylen och sedan kommer allt att rulla på av sig själv, och han ger också en förklaring på kemispråk till hur det kan ha gått till. Eftersom det är få läsare som är insatta i kemi, och ännu färre som har perspektiv på hur celler fungerar, så risken överhängande att han lyckas med sitt uppsåt. Det här är nämligen ingenting annat än ett sagoberättande i vetenskaplig förklädnad. Varför då?
Scenariot bygger på att 1. den aktuella proteinmolekylen och 2. den ljuskänsliga molekylen (cis-retinal) redan finns på plats och att proteinet är kopplat till 3. ett befintligt signalnätverk i en cell. Det här är mycket sofistikerade saker:
1. Det så kallade G-proteinet är en grupp av totalt 700-800 aminosyror i en specifik sekvens fördelade på tre sammankopplade underavdelningar (ett så kallat komplex).
2. Cis-retinal är en molekyl som aldrig påvisas genom något livets-spontana-uppkomst-experiment, utan kräver komplexa enzymsystem för att kunna tillverkas. Den går att framställa men då bara med hjälp av avancerad målstyrd kemiteknik (läs: design!).
3. Biologiska signalnätverk består av fyra delar: 1. En sensor (det som E. hävdar att det räcker med två mutationer för att skapa). 2. Transduktion som omvandlar och överför signalen på ett lämpligt sätt. 3. Integration som är en algoritm för att välja en lämplig åtgärd bland olika alternativ, och 4. en effektor som verkställer den valda åtgärden.
Allt detta behöver finnas på plats för att E.s mutationstrolleri ska fungera. Om något av det saknas spelar det ingen roll om en molekyl reagerar på ljus. Då ”förstår” ju inte cellen vad det är för mening med det.
Sedan finns det ytterligare en ”detalj” som E. väljer att inte berätta för läsarna:
Det räcker nämligen inte att ljus träffar den ljuskänsliga cis-retinalmolekylen och att det resulterar i en kaskad av ändamålsenliga kemiska reaktioner. När den belyses omvandlas den nämligen till en energifattigare variant (trans-retinal). Detta måste regenereras (återbildas) så att molekylen kan upprepa bedriften gång på gång, annars blir det en enstaka impuls och inget mer.
Evolutionsscenariot påminner om det i Rube Goldbergs skämtteckning här nedanför. Cigarettändaren (J) tänder stubintråden och raketen (K) far iväg med följden att linan (M) spänns och i förlängningen till att mannens mun torkas av. För att den här proceduren ska kunna upprepas en gång till behöver den förbrukade raketen ersättas med en ny. Raketen motsvarar förstås synsystemets cis-retinal.

”Självtorkande servett” av Rube Goldberg, Public domain, via Wikimedia Commons
Om någon tycker att maskinen verkar lite långsökt, så är det ingenting mot E.s scenario. För återbildningen av cis-retinal krävs åtminstone fem olika specifika enzymmolekyler bestående av sammanlagt 1100 aminosyror. (För övrigt tycker jag att Goldbergmaskiner är en träffande illustration av evolutionära berättelser överlag).
Till skillnad från synsystemet skulle servettorkningsmaskinen fungera lite halvbra i praktiken, men ingen skulle för den skull påstå att den inte var designad (av Goldberg).
Synsystemet är alltså ofantligt mycket mer komplext än E. vill berätta. Som tidigare påpekats: det krävs en kortlek innan man kan börja spela kort. Påståendet att det bara skulle krävas två ynka mutationer för att skapa synförmåga är alltså en sanning med sådan modifikation att det i praktiken är en ren lögn.
Om E. läser detta (vilket jag förutsätter) kommer han att utbrista: - Schmidt förstår ingenting, det är ju just det jag försöker förklara – evolutionen bygger på det som redan finns och behöver aldrig skapa någonting nytt.
Men "There is no such thing as a free lunch".37
Mina tankar går osökt till 1600-talets ”homunculi”. När den nederländske Nicolaas Hartsoekers studerade mänskliga spermier i sitt mikroskop tyckte han sig se att det satt en liten miniatyrmänniska inuti varje spermiehuvud. Man tänkte sig alltså på den tiden att det bara var kvantitativa skillnader mellan spermien och den vuxna människan. Det finns onekligen paralleller till E.s perspektiv.

Dessutom kommer jag att tänka på boken om Baron von Münchhausens fantastiska äventyr. Där kan vi läsa om den där dagen då baronen var ute och red på sin häst och råkade hamna mitt ute i ett träsk. Hästen och baronen sjönk allt djupare i dyn till dess att han i sin desperation kom på en lysande idé: han tog av sig hatten och tog ett rejält tag i sin egen kalufs, klämde ihop knäna och lyfte både sig själv och hästen upp ur dyn. Beslutsamt gjort kan man ju konstatera.
Vi vet att det är en skröna eftersom fysikens lagar inte tillåter sådant.
Vad var det nu E. skrev…:
ett fullt rimligt scenario.
E. är så angelägen om att visa att design är en överflödig hypotes att han inte uppmärksammar att han kombinerar filosofisk reduktionism med rent sagoberättande i vetenskaplig förklädnad och helt utan evidens från kemins område.
För att ytterliga understryka att evolutionen fungerar alldeles utmärkt utan att någonting nytt behöver introduceras ägnar E. ett avsnitt åt den så kallade endosymbiontteorin (s 61). Enligt denna bildades eukaryota celler och växtceller genom att en enklare cell invaderades av bakterier. Bakterierna avvecklades sedan till mitokondrier respektive kloroplaster. Jag tar inte utrymme till att kommentera saken närmare här, utan nöjer mig att konstatera att det här långtifrån är något bevisat faktum, även om det brukar framställas så för våra skolelever.
Endosymbiosteorin är populär, inte på grund av någon empirisk evidens, utan för att det saknas evolutionära alternativ. Berättelsen låter trovärdig i all dess enkelhet – en bakterie sväljer en annan bakterie och sedan är saken klar. Men faktum kvarstår – kontrasten mellan prokaryoter och eukaryoter är enorm och endosymbiontteorin skapar i praktiken fler problem än den löser.38 Ingen gratislunch för E. här heller alltså.
Under rubriken om fitness poängterar E. (s 62) att evolutionen handlar om reproduktionsframgång och ingenting annat. Hur komplex en organism är eller hur gammal den blir kvittar. Det som spelar roll är hur effektiv den är på att föra sina gener vidare till nästa generation, det vill säga hur många ungar den får. Många evolutionsbiologer jämför det här med cancer. Det som skiljer en cancercell från en frisk cell är att en så kallad proto-onkogen muterat. Exempelvis har den så kallade ras-genen i en frisk cell en funktion som kan liknad vid en gaspedal. En mutation kan leda till att gasen hänger sig i nedtryckt läge vilket leder till att cellen börjar dela sig okontrollerat; proto-onkogenen har omvandlats till en onkogen (cancercell). Den snabbare förökningstakten hos cancercellen ger den en konkurrensfördel på motsvarande sätt som en organism med högre reproduktiv framgång än sina konkurrenter. Vad innebär det här? Det innebär att cancercellerna till slut tar över organismen ända tills den dör.
Likheten med det naturliga urvalet är talande. En tumör av cancerceller består av celler som bär en nedbrytande mutation som gett dem en selektionsfördel i form av ökad fitness. Det är precis samma förhållande som råder i en population med organismer som är bärare av en fördelaktig mutation med en nedbrytande effekt, likt sicklecellanemigenen.
Om den evolutionära hypotesen om reproduktiv framgång som drivkraft för evolutionen vore sann, så skulle vi ha en evolution mot några få, och kanske till slut en enda, superorganism som likt cancercellerna konkurrerar ut alla andra celltyper. Den biologiska världen skulle se helt annorlunda ut om den historien vore sann. Den ekologiska dynamik och balans som vi kan observera i naturen säger oss att sanningen om selektionen är en annan. Det är tur. Om evolutionsteorin stämde skulle vårt öde vara beseglat precis som för den cancerdrabbade kroppen.
Ingen riktig skotte
Det finns inte jättemycket att säga om de avslutande delarna av bokens första avdelning. E. tar upp en av mekanismerna för artbildning som är helt okontroversiell. Jag nöjer mig med att kommentera några saker som E.:s påstår under rubriken makroevolution:
Ett exempel [på dramatiska förändringar i miljön som gör att nya utvecklingsvägar öppnas] är den katastrof som utrotade dinosaurierna för 65 miljoner år sedan. Knappt 20 miljoner år därefter finner vi fossil som representerar de flesta av dagens däggdjursordningar: gnagare, kaniner, fladdermöss, hovdjur, rovdjur, primater med flera. Det är ingen slump. Katastrofen innebar att många ekologiska nischer blev lediga, och det blev utrymme för däggdjuren som inte längre behövde konkurrera med dinosaurierna. (s 64)
E.s tes är den vanliga evolutionära berättelsen att en naturkatastrof (vanligen ett meteoritnedslag på Yucatanhalvön i nuvarande Mexico) utplånade dinosaurierna och att den drastiska miljöförändringen som det innebar möjliggjorde för de olika däggdjursordningarna att utvecklas, nu när de inte längre behövde konkurrera med dinosaurierna.
Stämmer berättelsen med evidensen? Nja, så där.
Det kan mycket väl vara så att de olika däggdjursfamiljerna var förhållandevis små till numerären när dinosaurierna hade sin storhetstid. Men det är mindre viktigt ur evolutionssynpunkt. E. ger läsaren samma intryck som man får via muséer, läromedel, högskoleutbildningar och populärvetenskap, nämligen att det från dinosauriernas ”era” bara återfinns lämningar av enstaka däggdjur och att såväl däggdjur som fåglar utvecklats ur just kräldjursförfäder från denna ”epok”.
Men det visar sig att 43 olika däggdjursarter finns representerade i en enda formation ”redan” i juralagren. Och i dinosaurieförande lager överlag har man påträffat representanter från närmare 300 släkten av däggdjur (många gånger flera arter inom enskilda släkten) vilket motsvarar 20-25 procent av alla nutida släkten.
Det finns rapporter om fossila fynd i krita (lagren med de rikaste dinosauriefynden) av till exempel ett indiskt hovdjur (Science 2007-11-09), igelkott (Victoria museum, Australien), opossum (South Australian museum) och av "stora mesozoiska däggdjur som levde av unga dinosaurier" (Nature 2005-01-13), och dessutom en kinesisk bäver från juralager (AAAS 2006-02-24). Och vad gäller fladdermössen så vet vi att den första (i bemärkelsen djupast begravda och påträffade) fossila fladdermusen, Icaronycteris, hade samma inbyggda system för ekolokalisering som nutida släktingar har. Den påträffas i och för sig i något högre lager än krita (tidig eocen), men en evolutionsbiolog skulle knappast mena att det tog en kvart för så sofistikerade system att evolvera. Även evolutionister anser därför att det fanns fladddermöss med åtminstone någon form av ekolokalisering redan i krita.
Det torde alltså inte vara någon tvekan om att det i samma sediment som vi återfinner dinosauriefossil också påträffar en mångfald av däggdjur. Sådana ”måste” klassas som primitiva, eftersom de inte stämmer överens med den evolutionära berättelsen. Och det skulle säkert vara E.s replik till det jag skriver. Min återreplik i så fall är att E. gör sig skyldig till det klassiska logiska felslutet som filosofen Antony Flew40 en gång myntade:
- Ingen skotte strör socker på gröten.- Men min vän Angus är skotte och gillar socker på sin gröt.
- Ah, jovisst, men ingen riktig skotte skulle strö socker på gröten.
Men hur tänker då bibeltroende biologer om det faktum att de däggdjur som återfinns bland dinosaurierna åtminstone delvis hade en annan, mer ”primitiv” anatomi än de nutida?
Som vi nyss konstaterade leder selektion och artbildning över tid till genetisk utarmning och specialisering. Det innebär samtidigt att tidiga former av organismer hade en större genetisk mångfald och var mer generella i fråga om sin anatomi än de nutida, vilket i evolutionisters ögon tolkas som ”primitiva drag” . Det kan illustreras av det ovan nämnda vargexemplet: Om vi skulle skapa en hybridform av alla de specialiserade hundraser som existerar idag skulle vi få tillbaka ett hunddjur som påminner om urhunden (sannolikt vargliknande). Vargliknande arter har ju mer generella och därmed skenbart mer primitiva drag än nutida hundraser.
De representanter av de olika däggdjursfamiljerna (och även familjer inom andra klasser bland ryggradsdjuren) som omkom i samband med den globala översvämningen på Noas tid (som Bibeln dokumenterar i 1 Mos 6-9) hade mer generella drag än nutidens representanter. Sedan dess har det skett en snabb specialisering och artbildning inom ramen för de skapade grundtyperna.41
Just den här frågan var en av de som behandlades vid Nordic Creation Research Conference i Malmö hösten 2024. I noten42 finns en länk till ett föredrag i ämnet av biologiprofessor Nigel Crompton från Cornerstone-universitetet i Michigan, USA. Föredraget finns även i skriftlig form i den Proceedings från konferensen som kommer att publiceras under våren 2026.43
E. konstaterar att:
Evolutionsteorin har visat sig vara en teori med stor förklaringskraft. (s 65)
Man kan se det på olika sätt. Vi har tidigare sett att det finns tveksamheter om huruvida evolutionsteorin är falsifierbar eller inte. Skulle så inte vara fallet är det lika korrekt att säga att en teori som likt den om evolutionen förklarar allt, samtidigt förklarar – ingenting.
E. presenterar ingen som helst evidens för att nya gener för nya egenskaper kan uppkomma. Hans enda argument för det är en tro eller förhoppning om att det kan ske genom gradvisa förändringar av till exempel enzymer. För att vara trovärdig skulle han behöva förklara det faktum att 10-30% av alla organismers genom utgörs av så kallade ”föräldralösa gener” (Orphan Genes, OG:s), även kallade ”taxonomiskt begränsade gener” som är unika för just den organismen. I människans fall handlar det om ett par hundra sådana. Någon känd evolutionär mekanism som kan förklara det existerar inte.
En brasklapp
Innan vi går vidare till nästa kapitel vill jag understryka att allting med evolutionsteorin inte är felaktigt, vilket meningsmotståndare till kreationism och ID brukar beskylla oss för att tro. Populationer av levande varelser förändras utan tvekan genom faktorer som genetisk drift, mutationer och selektion. Förändringarna över tid är emellertid alltid begränsade till de kategorier som organismerna tillhör. I fråga om ryggradsdjur innebär det vanligen den systematiska familjenivån (som hunddjur, kattdjur etc). Det här handlar uteslutande om så kallade mikroevolutionära förändringar. I fråga om den sortens förändringar är jag och E. relativt överens. Skiljelinjen mellan oss handlar om huruvida förändringar av levande varelser över tid är totalt obegränsade (vilket han hävdar), eller om de är begränsade till att ske inom ramarna för varje enskild (skapad) kategori av organismer, vilket är vår hållning som kreationister. På den punkten skulle en ID-supporter vara mindre tydlig (se kap 28 nedan)
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 9. Darwinism, vad är det?
E. skriver att han helst vill undvika ordet darwinism (s 66), med hänvisning dels till att ändelsen -ism associerar till en ideologi och dels till att det är oklart om det anspelar på Darwins syn eller moderna biologers.
(I det första avseendet kan jag erkänna att jag har en analog hållning när det gäller kreationism. Jag föredrar att säga ”kreationära förklaringar” än kreationistiska. Men det ska ses som en parentes).
E. tar kraftigt avstånd från idén att nazisttidens socialdarwinistiska människosyn med rashygien och utrotningsläger skulle ha någonting med evolutionsteorin att göra. Sådant där, menar han, beror på att man blandat ihop naturligt urval med artificiell selektion:
artificiell selektion, … inte något som går att härleda ur evolutionsteorin. (s 69)
Socialdarwinismen hade just ingenting med Darwin att göra. (s 69)
I egenskap av ID-anhängare och kreationister är vi eniga med E. om att det är djupt ovetenskapligt att påstå att en viss etnisk människogrupp skulle vara genetiskt överlägsen någon annan. Det vore som om en pudel reste sig upp och proklamerade sig som genetiskt ädlare än vargen. Vi har ju sett att det omvända råder.
Skillnaden är att E. knappast kan förankra sin syn i sin evolutionära världsbild. Som bibliska kreationister konstaterar vi att hela människosläktet härstammar från en enda familj (Noas) för omkring 4300 år sedan och att detta är i full harmoni med demografin.44 Men E. som har ett hundratals gånger längre perspektiv på mänskligheten kan inte alls med samma säkerhet dra samma slutsats. I så fall skulle han behöva förklara varför man kan ta för givet att en gren i det evolutionära trädet som utvecklats under hundratusentals år utan kontakt med andra grenar inte skulle kunna ha utvecklats till en genetisk nivå som över- eller understiger en annan grens.
Märk väl att jag inte för en sekund antyder att E. skulle vara rasist. Tvärt om – jag ser med tillfredsställelse på att vi har samsyn i en så viktig samhällsfråga. Men jag anser att hans korrekta syn inte kan motiveras på ett fullgott sätt utifrån hans evolutionära syn på verkligheten.
Historierevisionism
Men det finns någonting annat som är betydligt mer beklagansvärt i E.s resonemang: historierevisionism. I sin iver att försvara evolutionsteorin förnekar han nämligen evolutionsteorins uppenbara betydelse för nazitidens dårskap.
I början av 1900-talet var Ernst Haeckel (1834-1919) en av den tyska naturvetenskapens frontfigurer. Han var en hängiven darwinist. Han publicerade 1901 stamträdet över människans evolution så som man på den tiden såg på saken. Det såg ut så här:
”Natürliche SchöpfungsGeschichte”, Ernst Haeckel, Georg Reimer Verlag, Berlin 1911, s 743.
Det var en sådan syn på människan som de tyska barnen fick lära sig i skolan. Någon procent av dem hette Adolf i förnamn. Efter 1945 gör praktiskt taget ingen det längre, och vi vet alla anledningen. Tror du som läsare att det spelar någon roll för vuxenlivets värderingar vilken syn på människan som skolan presenterar som vetenskaplig sanning? Jag tror det. Och i fallet Adolf H är det ingen tvekan att hans evolutionära socialdarwinistiska människosyn påverkade det tyska samhälle som han var med och formade. E. hävdar att Hitler hade missuppfattat alltsammans, att
artificiell selektion … inte … går att härleda ur evolutionsteorin. (s 69)
Ursäkta ordvalet, men: Struntprat! Det är ett obestridligt historiskt faktum att ett av Darwins huvudargument var just den artificiella (mänskliga) selektionen av olika slag av husdjur som prototyp för evolutionen i den fria naturen. Och det var samma typ av resonemang som praktiserades genom olika former av rashygieniska ”åtgärder” under 1930-talet (och även senare). Människosynen spelar roll! Att som E. förneka darwinismens roll i det här avseendet är inte bara historierevisionism, det är dessutom precis samma logiska felslut som nyss: ”Ingen riktig skotte”. Det var ju selektionen som princip som nazisterna byggde sin politik på, inte bara den artificiella varianten av den!
↩ Tillbaka till innehållsförteckningen
![]()
I det här avslutande kapitlet i avdelning ett presenterar E. lite summariskt ungjordskreationism, gammaljordskreationism och teistisk evolutionism (TE).
På den dryga halvsida (s 71) där han berättar om föreningen Genesis och våra engelskspråkiga motsvarigheter AiG, CMI och ICR finns, förutom ett par smärre sakfel,45 en ganska grov desinformation. Han skriver att föreningen Genesis sprider skapelsetron
genom att kritisera framför allt evolutionsbiologi, paleontologi och geologi. (s 71)
– ett påstående som han dessutom upprepar längre fram i boken (s 83). Sanningen är att positiv evidens för design/skapelse är väl så viktigt som evolutionskritik för oss i föreningen.46
E.s beskrivning av teistisk evolutionism är betydligt fylligare och generellt mer positiv. Det är inte så konstigt, eftersom den vanligaste varianten av den här skapelsesynen köper evolutionsteorin helt reservationslöst. Han refererar (s 77) till den amerikanske vetenskapsfilosofen och teologen Ian Barbour som presenterat fyra olika sätt för religioner att förhålla sig till vetenskap:
1. Konflikt (”kreationistisk hållning”)
2. Självständighet (den vanligaste hållningen inom TE att religionen bör sköta sitt och vetenskapen sitt utan att överlappa varandras områden)3. Dialog (att företrädare för vetenskap och religion lär av, och berikar, varandra)
4. Integration (att vetenskap och religion ska integreras, ”central inom ID-rörelsen”)
E. förordar nr 2. Var och en bör sköta sitt och låta den andre vara i fred. Jag återkommer till det här under nästa rubrik och mot slutet av mitt dokument (kap 28). Han är förhållandevis neutral till nr 3 och starkt emot nr 1 och 4. Hans summering är:
Religionen duger inte som vetenskap, vetenskapen duger inte som religion. (s 76)
Det är svårt att ha någon invändning mot den formuleringen. Men E.s utgångspunkt är att den Gud som religionen representerar inte har någonting med den här fysiska världen att göra. Och där går naturligtvis våra meningar isär. Det har nämligen Bibelns Gud i allra högsta grad!
Avvisa teistisk evolution
Det finns flera skäl till att kristna bestämt bör avvisa teistisk evolution (TE):47
• Den främsta anledningen är att evolutionens mekanismer inte fungerar av skäl som framgår i det här dokumentet.
• Den har heller inte stöd i det fossila arkivet.48 • Företrädare för TE förkastar som regel den traditionella kristna läran om skapelsens ursprungliga godhet liksom ett historiskt syndafall med långtgående konsekvenser för skapelsen, i strid med Nya testamentets lära. • När det gäller skapelseprocessen menar somliga TE-anhängare att Gud inte hade kontroll över skapelseprocessen, han bara satte igång den. Andra menar att Gud utifrån sitt perspektiv har kontroll över den, medan den från människans perspektiv är fördold, gömd bakom mutationer och selektion. Det skulle innebära att Gud inte skulle vara synlig genom sina verk, vilket tydligt strider mot bibelsammanhang som Ps 19 och Rom 1:20.↩ Tillbaka till innehållsförteckningen
![]()
AVDELNING II
Kapitel 11. Intelligent design
Boken andra avdelning behandlar argumenten för intelligent design. Först gör E. en introduktion av vad han anser att intelligent design är, och därefter följer hans vederläggning av tretton ID-argument.
Jag har nu för avsikt att gå igenom dem i tur och ordning och analysera E.s argument för att de inte håller måttet.
E. har som vi redan konstaterade inledningsvis uppfattningen att ID är ovetenskaplig. Han skriver:
För att kunna förklara något fenomen vetenskapligt måste man ha sådan kunskap om den förmodade orsaken att man säkert kan knyta denna orsak till den förmodade verkan man studerar. (s 82)
Men det gör man inte enligt ID-rörelsen, menar han.
Det kommer att visa sig att E. har en dubbel standard i fråga om det här.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 12. Första argumentet: Vetenskapsargumentet
Det E. kallar vetenskapsargumentet är ID-rörelsens anspråk på att bedriva vetenskap genom så kallad ”design-detektering”. Han tycker emellertid inte att det är något relevant anspråk och skriver att:
ID … drar slutsatsen om design utan kunskap om designern, som det verkar rent intuitivt. (s 82)
Designdetektering – vad är det?
För dig som inte hört talas om designdetektering så ska jag först förklara vad som menas med begreppet. Det är då viktigt att veta att det finns ett angränsande begrepp som hör nära samman med design, och det är avsikt. När man säger att någonting är designat så är det liktydigt med att det finns en bakomliggande avsikt.
Biologen Michael Behe, som vi kommer att återkomma till längre fram, menar49 att man kan detektera design genom att observera ett ändamålsenligt arrangemang av delar (ljud, ord, kugghjul, logistiken i en överraskningsfest). Endast ett personligt intellekt (”mind”) kan åstadkomma sådana, och därför utgör det positiv evidens för design.
Inom en rad olika vetenskaper använder man sig rutinmässigt av metoder för att avgöra om det finns underliggande avsikter (design). Några exempel:
• Inom forensiska (kriminaltekniska) vetenskaper vill man ha svar på frågor som: Var det en olycka eller en avsiktlig skada?
• Inom kryptografi vill man ta reda på om felet i programvaran uppstod av en slump eller om det gjordes det med avsikt.• Inom arkeologin vill man veta om mönstret i stenarnas placering är slumpmässigt eller om de blev avsiktligt utplacerade.
• Inom AI-teknologin används Turing-testet för att besvara frågan: Är rapporten författad av en människa eller en dator-bot?
• Som lärare ställs man numera ständigt inför frågan: Är formuleringen elevens egen eller är det ett plagiat? Och så vidare.
När det gäller den sortens frågeställningar har E. säkert inga invändningar, men han avvisar bestämt varje försök att tillämpa dem på företeelser i naturen. Han har två argument för detta:
1. Dels att ID bygger vetenskapsargumentet på intuition (se citatet ovan).
2. Dels att det är bedrägligt av ID att särskilja mellan att påvisa design och att argumentera och spekulera om designerns egenskaper. Påstår man att någonting är resultatet av en design så måste man ha kunskap om designern, hävdar han (s 82). Och följaktligen är ID ovetenskaplig eftersom det blandar in egenskaperna hos en övernaturlig agent.
Det första argumentet visar att E. helt har missförstått Dembski. Alla människor (inklusive E.) kan omedelbart, via sin intuition, avgöra när företeelser har formats med avsikt. Det kan handla om så vitt skilda saker som konstverk, kommunikationssatelliter och texten du just läser.50
Just på grund av att intuitionen som sådan är alltför diffus för att använda vetenskapligt (magkänsla är ju knappast grund för en fällande dom) har Dembski och andra inom ID-nätverket liksom forskare inom diverse andra vetenskaper, arbetat fram kriterier och matematiska modeller för att göra design empiriskt påvisbar. Det handlar om att ta fram tekniker för att kunna urskilja intelligenta signaler mot en bakgrund av naturligt brus. Vi återkommer till det.
Det lätt att visa att inte heller E.s andra argumentet håller. I alla de fem exemplen i punkterna ovan är det inte det primära att få reda på de personliga egenskaperna hos den eller de personer som varit aktiva (agenterna), utan bara om det är ett resultat av rena tillfälligheter eller inte.
E. tycker att det är just referensen till Gud som är det stora problemet. Men William Dembski tog effektivt udden av det argumentet i sin klassiska bok Intelligent Design51. Han skriver:
Stoikerna använde sig flitigt av begreppet design i sina kosmologier, även om de inte hade någon uppfattning om en transcendent, än mindre en personlig, mirakelverkande Gud. (Intelligent Design s 88, förf. övers).
De här argumenten för att förklara ID ovetenskaplig är alltså ohållbara, och som jag nämnde i all hast leder de dessutom till att E. tvingas att tillämpa en dubbel standard. Han skriver:
Analogin haltar. För att en forskare ska ta till en ”agentförklaring” krävs att man vet att det faktiskt existerar en möjlig agent/designer, och att man vet att agentens sätt att arbeta kan förklara det fenomen man studerar. Man förklarar alltså fenomen med faktorer man vet någonting om. (s 84)allt blir rena gissningar (s 85)
Vetenskapliga förklaringar bygger på att den kunskap man har om en viss ’agent’ kan förklara ett fenomen. I en ID-förklaring har man emellertid ingen kunskap om sin ’agent’, inte ens om vederbörande existerar. Man uppfinner helt enkelt en agent som man antar har sådana egenskaper att det helt kan förklara fenomenet man studerar. (s 87-88)
Det här är ett argument från okunnigheten. (s 86)
I nästa andetag argumenterar E. för att den så kallade SETI52-forskningen ryms helt inom ramen för vetenskapen. Central i denna forskning är analyser av radiovågor från rymden med hjälp av radioteleskop. Man söker efter mönster i radiovågornas brus som skulle kunna indikera intelligenta avsändare, därför att det är vad man förväntar sig. I den klassiska boken Contact53 av astronomen Carl Sagan (1985) fångar radioteleskop i Mexico upp en annorlunda signal i form av en sekvens av de 261 första primtalen.54 Eftersom det inte finns någon känd naturlig process som skulle kunna generera en primtalsserie så drar man slutsatsen att det är intelligenta utomjordningar som är avsändare. Följaktligen designdetektion i allra högsta grad. I det här fallet science fiction, men samtidigt en fullt vedertagen vetenskaplig(!) metod inom SETI.
Låt oss jämföra det här med de ”kriterier” som E. nämner i de båda citaten. Han menar att en förutsättning för en ”agentförklaring” är
man vet att det faktiskt existerar en möjlig agent/designer. (s 84)
Men vet man inom SETI att det verkligen finns utomjordisk intelligens? Nej, det är en obevisad hypotes, inte mer än så.
En annan förutsättning menar E. är
att man vet att agentens sätt att arbeta kan förklara det fenomen man studerar. (s 84)
Vet man inom SETI att eventuella utomjordiska intelligenser har koll på primtal? Tja, man kanske skulle kunna förmoda det, men vi kan knappast veta, eller hur?
Man förklarar alltså fenomen med faktorer man vet någonting om (s 84)
Men vad vet vi egentligen om rymdvarelser?
I en ID-förklaring har man emellertid ingen kunskap om sin ’agent’, inte ens om vederbörande existerar. Man uppfinner helt enkelt en agent som man antar har sådana egenskaper att det helt kan förklara fenomenet man studerar. (s 88)Det här är ett argument från okunnigheten. (s 86)
När Isaac Newton lade fram sin gravitationsteori och föreslog att himlakropparna attraherade varandra på väldiga avstånd så var det närmast en magisk kraft han hänvisade till och införde i sina ekvationer. Han förklarade alltså fenomen (himlakropparnas rörelser) med en faktor (gravitationskraften) vars natur man inte visste någonting om (och knappt vet än idag). Tänk om man följt en dåtida E.s råd och förklarat det ovetenskapligt!
Dessutom … skulle inte just det här vara precis lika tillämpbart på de utomjordingar som man söker efter inom SETI-projektet? Man ”uppfinner” ju onekligen en utomjording som förväntas ha egenskapen att exempelvis producera primtal. Vad vet vi då om utomjordningar? Ingenting. Bara gissningar där också. Jag skulle rekommendera E. att kontakta forskarna vid SETI. och förklara för dem att de argumenterar ifrån okunnighet.
Det råder alltså ingen som helst tvekan om att det är hundra procent vetenskapligt att tillämpa designdetektion vid studiet av naturen, och vi behöver inte alls spekulera om agentens egenskaper och natur.
Det finns faktiskt ett svenskt SETI-projekt som går under namnet Hephaistos55 vid institutionen för astronomi vid Uppsala universitet. Man studerar avlägsna galaxer i hopp om att kunna observera historiska förändringar av dem. I så fall är hypotesen att galaxerna bebos av så intelligenta varelser att de till och med förmår förändra galaxernas utseende och egenskaper. Nu börjar vi närma oss rent gudomliga nivåer av intelligens. Och allt helt inom ramen för etablerad vetenskap. Man kan förstås ställa sig frågan om hur mäktig en intelligent varelse behöver bli för att inte längre få rymmas inom materialismen.
Mot bakgrund av detta måste den naturliga slutsatsen bli att E. har ett annat incitament bakom det han skriver och att ovanstående mest är ett svepskäl. Synpunkten att det skulle vara ovetenskapligt att detektera design i naturen går alltså bevisligen inte att vidhålla utan att på samma gång förklara många områden inom fullt accepterad forskning för ovetenskapliga, vilket vi redan konstaterat.
Kropp och själ
Det här besannas också vid den fortsatta läsningen av kapitlet. Det visar sig nu att det är kravet på den stränga naturalismen som är E.s egentliga argument. Förutom att det är omöjligt att studera övernaturliga varelser med vanliga vetenskapliga metoder (vilket inte finns någon anledning att bestrida, men som man inom ID-rörelsen inte heller påstår) för han nu fram argumentet att det är fel att utgå från att intellekt, vare sig Guds som människors, skulle kunna existera oknutna till materia:
Men människans intelligens är … en egenskap hos hennes fysiska kropp. (s 87)Man kan alltså inte ge något trovärdigt svar på frågan om hur en okroppslig intelligens kan styra processer i den fysiska verkligheten. (s 87)
Nu uttalar sig biologen E. om saker som ligger utanför hans kompetensområde. Frågan om själens och medvetandets existens och natur är framför allt en filosofisk och religiös fråga. Lika lite som E. kan veta att hans medvetande upphör i samma ögonblick som hans centrala nervsystem upphör att fungera, lika lite kan han uttala sig med någon vetenskaplig trovärdighet om Guds natur och existens.
Visst får E. ha sin tro och uppfattning i fråga om detta. Men majoriteten av jordens befolkning tycker annorlunda. ID:s och kreationismens förhållningssätt är att icke-materiella, personliga och medvetna intelligenser – det vi i dagligt tal kallar ”själar” – ständigt växelverkar med den materiella världen genom allting vi säger och gör. Bevisbördan vilar på E. att visa att vi har fel.
Själv får jag nog säga att det finns gott om evidens i samband med nära-döden-upplevelser som talar emot Erkells syn på det här området. Jag minns också en anekdot av tidigare omnämnde Antony Flew från hans bok There is no a God (se not 38).
Det är en fiktiv berättelse om två ociviliserade infödingar på en avlägsen strand som råkat hitta en mobiltelefon i sanden. När en av dem råkar trycka in en tangent hörs en röst som talar. Med skräckblandad förtjusning konstaterar han att det är någon som talar genom apparaten.
- Struntprat, säger den skeptiske infödingskamraten. Rösten måste vara ett resultat av hur de olika delarna är sammansatta, och jag kan bevisa att jag har rätt!
- Hur då?
- Ge den till mig, säger han och drämmer den med all kraft i en bergvägg. Rösten tystnar.
- Där ser du, konstaterar han med triumf i blicken, Det var som jag sa, rösten satt i delarna. De gick sönder och rösten försvann!
Tycker du att den skeptiske infödingens resonemang var övertygande? Troligtvis inte med vår tids kunskap om mobiltelefoni. Men infödingarna som saknade vårt perspektiv skulle säkerligen ha fortsatt diskussionen där borta i det avlägsna landet (som på något mystiskt sätt försetts med mobiltäckning).
Analogin med själen och medvetandet är nog ganska uppenbar. Det är naturligtvis så att när man blir tillräckligt gammal eller sjuk så dör man. Men frågan om själens existens kvarstår för den skull obesvarad, precis som röstens ursprung för infödingarna. Det var dit Antony Flew ville komma. Och jag med. Det duger inte att E. levererar den sortens argument för sin tes.
Frågan om förhållandet mellan kropp och själ har en rent fysikalisk motsvarighet. Det är den om varför materiella föremål (som till exempel stjärnor) lyder icke-materiella naturlagar (som gravitationslagen). Stjärnor åldras och slocknar, men det gör inte naturlagarna.
Dessutom finns den informationsteoretiska motsvarighet som vi kommer att behandla i kapitel 17. Den levande världens DNA-molekyler bär bevisligen meningsfull information i form av specificering av alla levande varelsers uppbyggnad och egenskaper, men informationen som sådan utgör ingen materiell del av DNA:s nukleotider, lika lite som informationen i den här texten utgör en inneboende del av varje enskild bokstav.
Slutsats: E.s argument mot IDs anspråk på vetenskap håller inte.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 13. Andra argumentet: Konstruktörsargumentet
Kreationister och ID-förespråkare pekar, liksom Bibelns författare, på att skapelsens olika strukturer vittnar om sin skapare.56 Inom teistisk evolutionism menar man däremot att så inte är fallet, design går inte att påvisa eftersom evolutionens gud agerat dold bakom naturliga mekanismer som mutationer och naturligt urval. Det är den teologin och guden som E. rekommenderar läsaren att anamma, 57 eftersom den lämnar naturvetenskapen och den materiella världen i fred, till skillnad från Bibelns Gud.
E. noterar helt riktigt att anledningen till att man inom ID argumenterar så, är att naturen vimlar av företeelser som vittnar om en designers ofattbara intelligens. Den slutsatsen är emellertid inte bara ett intuitivt eller emotionellt konstaterande, utan en slutsats baserad på att många strukturer har sådana egenskaper att de inte skulle kunna uppstå genom en gradvis händelsekedja. Vi har redan berört ämnet (kap 8) och skall också återkomma till det längre fram (kap 16).
E.s invändningar mot konstruktionsargumentet är dels det klassiska anti-kreationistiska Dawkins-argumentet att världen är full av exempel på ”dålig design”, eller som det ibland kallas: suboptimal design, samt att det förutsätter att designern ”tänker som vi som vi själva gör.” (s 92).
Sett utifrån ett kreationistiskt perspektiv är det där förstås ingen överraskning. Eftersom vi är skapade av Gud, och enligt Bibeln uttryckligen skapade till hans avbilder, så är det helt förväntat att Designern arbetade efter liknande principer som intelligenta konstruktörer och programmerare gör idag. Möjligen kan man säga att det inte är lika lätt att motivera ur ett renodlat ID-perspektiv.
Enligt E. är alla exempel på misslyckad design och utdöda djurarter i det förflutna precis vad en sekulär biolog skulle förvänta sig av en planlös evolution som släpar med sig spåren efter sin evolutionära historia.(s 92) Som exempel tar han upp den trånga födelsekanalen (s 89) och menstruationsblödningar hos kvinnor (s 90), placeringen av mannens testiklar (s 90) och prostatakörtelns konstruktion (s 90).
Underoptimal design – en självmotsägelse
Redan inledningsvis kan vi konstatera att det är en självmotsägelse att ens använda begreppet underoptimal design som ett argument för frånvaro av design. Det går nämligen knappast att hitta någon definition av design som inte är ett resultat av en intelligent aktivitet. Wikipedia definierar exempelvis begreppet så här:
Formgivning/design är den ändamålsenliga utformningen genom en formgivare/designer av eller framställda produkter och miljöer.
Byter vi ut orden ”ändamålsenliga” mot ”planlösa” och ”genom en” mot ”utan någon” så blir hela definitionen obegriplig och fullständigt irrelevant. Vid all design/formgivning är nämligen just ändamålsenlighet, planering och förutseende centrala ingredienser som förutsätter intelligenta aktörer, detta alldeles oavsett om vi tycker att föremålen i fråga passar vår smak eller inte.
Optimal design – vad är det?
Vad menar vi egentligen med en optimal design? Kanske räcker det att fråga sig vilket som är det optimala skärverktyget i ett kök - en kökskniv eller en laser? Av samma skäl måste vi ifrågasätta idén att någonting är suboptimalt på grund av att det skulle gå att hitta en alternativ konstruktion som skulle kunna utföra uppgiften med större precision. I stället blir frågan om den aktuella funktionen kan utföras tillräckligt bra givet organismens behov i den miljö i vilken den lever.
Laptopen jag just nu skriver på är långt ifrån optimal i alla enskilda avseenden. Skärmen är ganska liten, hårddisken likaså, tangentbordet kunde vara lite mer ergonomiskt och så vidare. Men sammantaget utgör den en alldeles förträfflig designkompromiss som gör att jag inte alls känner mig frestad att byta ut den mot en stationär dator även om en sådan skulle kunna lagra betydligt mer data. Den bästa designen är inte optimal i sig, utan den utgörs av den optimala designkompromissen. Det är de optimala designkompromisserna som är den princip som präglar den levande världen. Vi kunde haft falkens ögon, men klarar oss alldeles utmärkt med de vi har.
Placeringen av mannens testiklar är en kompromiss mellan fördelen i ökad fertilitet på grund av att temperaturen vid spermieproduktionen kan hållas optimal, och nackdelen i ökad sårbarhet. E. borde även ur sitt evolutionistiska perspektiv inse att om ”evolutionen” gång på gång återkommit till samma konstruktion hos en mängd olika däggdjur så borde designkompromissen vara förhållandevis optimal.
Icke-optimal design kontra icke-design
E. angriper därför en halmdocka58 av det ID-argument han vill vederlägga. Jag vill tro att det beror på okunskap, men även det är klandervärt eftersom han gör anspråk på att veta vad han talar om.
ID-rörelsens företrädare, som till exempel William Dembski är noga med att betona att Intelligent design som begrepp måste särskiljas från å ena sidan skenbar design och å andra sidan optimal design. Låt oss ta ett konkret exempel:
Är allting på min gamla Ford Focus optimalt? Det kan man knappast säga. I princip alla komponenter skulle kunna vara effektivare. Dessutom är den sliten efter 20 år och 32 000 mil.
Men är den designad? Ja, det råder det naturligtvis inget som helst tvivel om. Designen är ingen illusion, det vill säga ett resultat av rena tillfälligheter, utan av en mängd yrkesgruppers samlade kompetens.
Det är den här betydelsen av design som ID menar är relevant och påvisbar. Den, menar man, återfinns på alla nivåer i naturen.
Jag nämnde tidigare om den engelske prästen och filosofen William Paleys (1743-1805) berömda urmakaranalogi. I sin analogi jämförde han planeternas regelbundna rörelser i solsystemet med ett fickur som någon hittar i naturen. I båda fallen, menade han, måste det finnas en intelligent formgivare.
Med den här analogin som bakgrund kan man säga att evolutionister som kritiserar skapelsetron med argumentet att levande varelser innehåller strukturer som de anser vara underoptimala egentligen säger att den där klockan i sanden kunde ha utformats på ett bättre sätt. Den kunde till exempel ha gått mer exakt, den kunde ha haft tydligare visare eller större siffror på urtavlan.
Men med den sortens påståenden missar de helt målet. Det frågan egentligen gäller är nämligen inte alls ifall klockan är optimal ur alla tänkbara aspekter, utan att det de facto ligger en klocka på stranden. Sol, vind, vatten och sand kan inte producera en klocka över huvud taget. Varför skulle tecken på en underoptimal design utgöra evidens för total frånvaro av design?
E. fokuserar däremot helt på de båda andra versionerna av design som är betydelselösa i sammanhanget. Optimal design förekommer inte i naturen, och knappast inom teknologin heller. Skenbar design kan förekomma, om än relativt sällsynt. Hans foto på en katt (s 164) illustrerar det (eller rättare sagt hade gjort det om det varit i färg). ID:s designkriterier har med avsikt formulerats så återhållsamt att den sortens falska designindikationer ska elimineras.
Hur kan vi veta att en viss design är underoptimal?
Att bedöma hur optimal en viss design i naturen är kräver både kunskap och perspektiv. Detta är nästintill omöjligt att uttala sig med säkerhet om. På 1970-talet när jag läste en kurs i molekylärbiologi på Chalmers var den allmänna känslan att det var ett område där man hade riktigt bra koll på läget. Hade man det? Knappast utifrån dagens utsiktspunkt, eller hur? Det kan finnas, eller rättare sagt – det finns säkerligen faktorer som vi missat, underskattat eller underlåtit att beakta och som gör att det som idag förefaller vara en suboptimal design mycket väl en dag kan visa sig vara optimal i ett helhetsperspektiv.
En annorlunda värld
Det skapelsekritiska argumentet om underoptimal design förutsätter också att den design vi idag ser hos en given organism är densamma som denna hade när den skapades av Gud. Men det är långt ifrån säkert, i synnerhet utifrån ett bibliskt perspektiv där Gud skapar en värld som är alltigenom ”god”, ja rentav ”mycket god”, men även inom ID-rörelsen för man en liknande argumentation. Bibeln vittnar om en värld som skapades fulländad (däremot inte med nödvändighet ”optimal” i den snäva bemärkelse som en evolutionist skulle hävda) men som på grund av människans historiska syndafall kom att präglas av sönderfall och död. När vi studerar designen hos nutida organismer och bedömer den vara suboptimal så kan det alltså mycket väl hända att det original som Gud en gång skapade var både ett och flera strån vassare.
Det är för övrigt lite kul att E.s väljer just kvinnans födelsekanal som sitt första exempel på dålig design, eftersom Bibeln uttryckligen säger att det är en direkt konsekvens av syndafallet och därmed något som inte var en del i Guds ursprungliga skapelse:
Till kvinnan sade han: Jag ska göra din smärta stor när du blir havande. Med smärta ska du föda dina barn. (1 Mos 3:17)
Prostatakörteln
Det finns funktionella skäl till prostatakörtelns konstruktion och placering. Om mäns prostataproblem berott på ett designfel skulle alla män ha besvär, men så är inte fallet. Prostatabesvär är relativt ovanligt bland japanska män som bor i Japan, till skillnad från japanska män bosatta i USA, vilket antyder att det är andra faktorer som kost och livsstil som inverkar. Konstruktionen är liknande hos hannarna hos många däggdjur. Om systemet varit dåligt borde väl E. ha förväntat sig att det naturliga urvalet skulle ha eliminerat det? Sedan kan man förstås tillägga att problem som likt prostata uppkommer på ålderns höst knappast är något argument mot design, lika lite som en rostig framskärm skulle vara något som talar mot att en bil skulle vara designad.
Förändrade miljöer
Evolutionsteorin förutsätter att det inte finns några gränser för den biologiska variationen – en mikrob kan utvecklas till en människa, bara tiden medger det. Bibeltroende biologer hänvisar i stället till den samlade erfarenheten från växtförädling och djuravel och hävdar motsatsen – variationen har gränser, nämligen de som separerar de av Gud skapade grundarterna från varandra. En bibliskt grundad hypotes är att många organismer skapades av Gud för att leva i vissa speciella miljöer (nischer) som inte längre existerar eftersom syndafallet som sådant och den följande syndafloden drastiskt kom att förändra livsvillkoren på jorden. Genetisk evidens tyder på att Gud i sitt förutseende skapade grundarterna med en förprogrammerad förmåga till anpassning i en föränderlig värld, men organismer som utsattes för de mest drastiska miljöförändringarna dog ut, som till exempel trilobiterna. Organismer som tvingades tangera gränsen för sin anpassningsförmåga för att överleva skulle kanske därför uppvisa en del underoptimala egenskaper idag, som till exempel jättepandans och koalornas extrema beroende av en enda födokälla (bambu respektive eukalyptus).
Icke-optimal design – en ekologisk nödvändighet
Det är helt avgörande för stabila ekosystem att populationerna av organismer balanserar varandra och att inte enskilda arter tar över, precis som är fallet med cellerna i en frisk kropp. En nödvändig förutsättning för det är just suboptimalitet, det vill säga att alla organismer har begränsningar i fråga om sina egenskaper. Faktorer som livslängd, förekomst av predatorer, populationsstorlek, antal avkomma, förekomsten av försvarsmekanismer varierar stort mellan olika organismer men är på totalen i balans i ett stabilt ekosystem. Massförökning som leder till väldiga populationer förekommer egentligen bara i kortvarigt i samband med kraftiga miljöstörningar. Det naturliga urvalets funktion i det här sammanhanget förefaller vara att upprätthålla den ekologiska balansen i ekosystemen snarare än att slipa fram nya strukturer, vilket naturligtvis hade förutsatt konstruktiva mutationer som lyser med sin frånvaro.
Underoptimal design som evolutionsargument
Designkritikerna hävdar alltså att underoptimal design är ett motargument till att livsformerna skapats av Gud. Samtidigt menar de att detta i stället är ett argument för en evolution av livet. Det där sista är emellertid en sanning med modifikation. Att naturen är full av strukturer av varierande ”optimalitet” är någonting som en evolutionist som E. förväntar sig eftersom den bygger på små förändringar som långsamt ackumuleras (ansamlas) över tid. I ett sådant scenario är det förväntat med en mängd överblivet skräp. Eftersom designföreträdare menar att designer skapade hela system med dess olika komponenter samtidigt finns ingen sådan förväntan hos dem/oss.
Ingen evolutionist skulle heller förneka att det finns en hel del saker i naturen som måste betraktas som nästintill optimala – det kan vi ofta höra programledarna förklara i de påkostade naturprogrammen på TV. Men har vi någonsin hört David Attenborough eller någon annan känd evolutionsförespråkare beskriva någon helt genial förmåga i djur- eller växtvärlden för att i nästa andetag konstatera att det där är ett argument mot evolutionsteorin? Knappast! Det vi i stället alltid får höra är i stället lovsången till det naturliga urvalet som under miljarder år lyckats finslipa alla detaljer intill fulländning. Återigen – slutsatsen kan bara bli en: evolutionsteorin är så anpassningsbar att den kan förklara precis allting – det perfekta såväl som det imperfekta. Det är teorins styrka men samtidigt dess fundamentala svaghet.
Den krassa sanningen är att underoptimal design aldrig betraktades som något evolutionsargument förrän det började användas i det här omskrivna sammanhanget. Det används av evolutionister enbart för att dessa anser att det motsäger existensen av en Skapare – vilket vi nu visat att det inte alls gör!
E.:s och andra evolutionisters tes är att allting i naturen som ser ut som om det vore avsiktligt designat bara är skenbar design. Att den antas ha uppkommit genom mutationer och naturligt urval (eller någon annan naturlig mekanism), och att ofullkomligheter och grymhet är helt förväntade egenskaper från sådana mekanismer. Problemet är att sådana mekanismer kan inte förklara de komplexa, informationsrika strukturerna i naturen som i stället utgör adelsmärket för en reell intelligent design.
Förekomsten av dålig design i naturen är inget argument för evolution, utan snarare ett problem. Varför har inte selektionen eliminerat djur så sårbara som jättepandan och koalan. Pandan som dessutom har stora reproduktionssvårigheter. Det borde E. ta sig en funderare på.
Evolutionistisk teologi?
Avslutningsvis finns det anledning att ifrågasätta hur pass relevant det är att ateistiska och agnostiska evolutionister oblygt för fram sina synpunkter på den Guds egenskaper och bevekelsegrunder som de i praktiken inte tror existerar. Det är nämligen vad de gör när de hävdar att den kristne guden skulle vara tvungen att skapa allt levande med en hyperoptimal design i varje enskild detalj. Finns det någon teologisk princip som skulle göra det nödvändigt? Inte mig veterligen.
Man borde ställa sig frågan: Hur skulle strukturen X hos organismen Y kunna förbättras under de betingelser den lever. I stället frågar man ”Vilken sorts Gud skulle skapa en sådan struktur?”.
Sådant understryker att personer som inte tror på Gud bör hålla sig utanför teologins område. Slutsats: Designargumentet håller – världen är verkligen skapad av Gud!
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 14. Tredje argumentet: Argumentet om ”skräp-DNA”
På mitt skrivbord ligger just nu ett utdrag från Dr Robert Wiedersheims bok Der Bau des Menschen als Zeugnis für seine Vergangenheit från 1902. Med visst utrymme för tolkning har jag kommit upp i 152 organ i människokroppen som Wiedersheim ansåg vara rudiment, alltså helt enkelt evolutionärt skräp.
Vi hittar inga sådana listor i nutida anatomiböcker. Av någon outgrundlig anledning har de senaste hundratjugofyra åren inneburit en makalös utveckling av funktioner hos hypofysen, binjurarna och mycket, mycket annat.
Eller?
Eller kan det möjligen vara så att det i stället är vetenskapen som gjort stora framsteg i att upptäcka funktioner i alla de strukturer som man vid 1900-talets början misstog för meningslösa slemsäckar?
Naturligtvis är det så. Men biologer tycks genom tiderna haft en tendens att skräp-förklara företeelser i naturen som man inte förstår. Varför då? Svaret torde vara uppenbart: det finns ingen som helst anledning för en evolutionsbiolog att förvänta sig att slumpmässiga mutationer och naturligt urval skulle kunna åstadkomma varelser där allting är funktionellt. Evolutionen är ju ingen målstyrd process likt de som designat bilar och datorer, där man har all rätt att förvänta sig en bakomliggande tanke och syfte med varje tangent och reglage.
En skräp-historik
DNA-molekylens struktur upptäcktes av Watson och Crick 1953. Ganska snart började man misstänka att bara en del av DNA kodade för proteiner, och vad var naturligare för evolutionsbiologer än att dra slutsatsen att resterande DNA var – just det – skräp. Så blev det. E. konstaterar också mycket riktigt att man redan i slutet av 50-talet börjat tänka i termer av genetiskt skräp. Det var emellertid först 1972 som termen ”junk DNA” började användas i den vetenskapliga litteraturen om eukaryota genom.59 Det användes då i bemärkelsen DNA som inte var utsatt för selektion (eftersom det ansågs sakna funktion att välja ut).
Under senare delen av 1970-talet kom uppfattningen om icke-kodande DNA som funktionslöst att dominera bland biologer och genetiker. Det rådde praktiskt taget total enighet under 1980- och 90-talen. När man på 80-talet upptäckte så kallade introner och transposoner så tycktes det bekräfta den här bilden. Först mot slutet av 1990-talet började man ana att vissa delar av ”skräpet” skulle kunna ha vissa funktioner (t.ex. regulatoriska områden).
I början av 2000-talet presenterades resultaten av HUGO-projektet – kartläggningen av människans genom. Det visade att bara 1-2 procent av det mänskliga genomet kodade för proteiner. Det förstärkte ytterligare uppfattningen att en stor del av arvsmassan bestod av icke-funktionellt DNA.
Under hela den här tidsperioden fanns det enstaka forskare som var öppna för möjligheten att det bland allt skräp skulle kunna dölja sig avsnitt av DNA som kunde vara funktionellt. Jag kan tycka att det vore märkligt annars, eftersom det vid den här tiden hade blivit uppenbart att den där listan över rudimentära organ hunnit krympa avsevärt – egentligen var ju ”skräp-DNA” bara ett nytt rudiment, fast den här gången på molekylnivå. Men, och det är huvudsaken, det råder ingen som helst tvekan om att ett enigt vetenskapssamfund höll merparten av vårt DNA för meningslöst skräp.
Men det fanns förstås en kategori vetenskapsmän som inte alls delade den uppfattningen. Det var kreationister, och så småningom även anhängare av intelligent design. Från deras utgångspunkt med en intelligent designer/skapare vore det ytterst oväntat om arvsmassan till största delen skulle vara ett skrotupplag, precis som deras föregångare under första halvan av 1900-talet hade opponerat sig mot Wiedersheims och andra evolutionisters åsikt att kroppen var full av skräp.
Det ovan sagda är den historiska beskrivningen av begreppet ”skräp-DNA” fram till 2012. Den läsare som känner tveksamhet kan snabbt få min berättelse bekräftad genom googling eller ChatGPT.
Historierevisionism – igen!
E. redogör också för den här historien (s 93-95). Men den bild han förmedlar till oss läsare är väldigt annorlunda. E. nämner (mycket riktigt) att det förekom en eller annan person under hela den här tidsperioden som hyste tvivel om att allt så kallat skräp-DNA saknade funktion. Men han underlåter att nämna om den massiva konsensus som rådde inom området under de 40 år som passerade från 1972 till 2012! Idén att skräp-DNA inte är skräp är inte en uppfattning som dykt upp tack vare evolutionsteorin utan trots den.
Här använder sig E. av samma taktik som självaste Richard Dawkins. I över trettio år hävdade denne berömde ateist att icke-proteinkodande DNA är parasitiskt skräp. I sin bok The selfish gene formade han flera generationers föreställningsvärld. Men när ENCODE-projektet (mer om det nedan) presenterade sina resultat 2012 vände han kappan efter vinden och menade plötsligt att det i själva verket var precis vad en darwinist förväntar sig – att hitta ändamålsenlighet i naturen. Det här, kära läsare, är varken mer eller mindre än historieförfalskning. Och återigen en indikation på att evolutionsteorin är oanvändbar – den förklarar allting och samtidigt ingenting.
Vad värre är: den evolutionära prediktionen om icke-proteinkodande DNA som skräp har hindrat och försenat den vetenskapliga utvecklingen på området i nästan ett halvt sekel!
ID-förespråkares och kreationisters prediktion är, och har alltid varit, att en hög men ospecificerad andel av arvsmassan har funktion och att det därför är av stort vetenskapligt intresse att studera denna del av arvsmassan för att upptäcka och dra nytta av resultaten!
Bortförklaringar
Historierevisionism har alltid ett dolt syfte. Vad kan det vara i det här fallet?
Saken är uppenbar. Det är djupt störande för en evolutionist att en förutsägelse utifrån ett skapelseperspektiv på naturen håller på att uppfyllas framför våra ögon och att ett halvsekellångt evolutionärt perspektiv håller på att grusas. Hur ska E. lyckas fördunkla detta?
Han gör det genom att angripa det på tre sätt. För det första hävdar han att ID förväntar sig att människans arvsmassa är 100-procentigt felfri (s 99). Det är nämligen lätt att visa att det inte stämmer. För det andra beskyller han forskarna inom ENCODE-projektet för att dra felaktiga slutsatser eftersom de tycks ge stöd åt ID-uppfattningen (s 98-99). Och för det tredje skriver han att även om ENCODE-forskarna och ID skulle få rätt, så skulle det inte spela någon roll eftersom det kvittar för evolutionsteorin om det finns något skräp i arvsmassan eller inte (s 99). Onekligen en smart helgardering av E. Låt oss behandla de tre invändningarna i tur och ordning:
Allt DNA funktionellt?
Först förväntningarna i fråga om det genetiska skräpet. E. skriver att ID-förespråkare hävdar att eftersom det finns en designer så
(1) borde det vara så att allt DNA är funktionellt. (s 93)
Evolutionsbiologer, däremot,
säger … inte mycket … om saken. (s 95)
Och:
(2) Påståendet att evolutionsteorin skulle kräva att vårt genom måste innehålla stora mängder skräp-DNA är alltså inte sant. (s 96)
Stämmer det här verkligen (1)? Hävdar ID (och för den skull kreationister) att allt DNA är funktionellt? Jag vet att E. inte kan hitta ett enda sådant påstående i vare sig litteratur eller videomaterial. Designförespråkare menar att de rådande naturkrafterna bidrar till att långsamt men säkert bryta ner skapelsen utifrån det ursprungliga optimala starttillståndet. I fråga om bibliska kreationister är det särskilt uppenbart eftersom man/vi hänvisar till ett historiskt syndafall med kosmiska konsekvenser, däribland mutationer som åstadkommer skador på det genetiska materialet (vilket ingen torde kunna förneka). Däremot gör ID ingen prediktion om någon specifik procentsats, mer än att funktionaliteten förväntas vara hög. Och alltid högre än som är konstaterat vid en viss given tidpunkt, eftersom det alltid går att upptäcka nya funktioner i tidigare förmodat skräp men sällan det omvända. Att ID och kreationister alltså är öppna för möjligheten att genomet ursprungligen var till 100 procent funktionellt innebär inte att det skulle vara det här och nu. Därför far E. återigen med osanning. ID menar inte att allt DNA är funktionellt.
Eftersom föreställningen om ett till hundra procent funktionellt DNA är grunden för E.s kritik skulle vi med all rätt kunna avfärda hela bokens kapitel 14 som irrelevant, men för den goda sakens skulle ska vi ändå analysera dess innehåll.
Men hans andra påstående då (2), det om skräp-DNA i det evolutionära perspektivet? stämmer det inte att evolutionister räknat med mycket DNA-skräp? Visst gör det! Det är exakt vad sekulära biologer förväntat sig under 40 års tid. Det går inte att finna något belägg för E.s påstående att ett i huvudsak funktionellt DNA skulle vara någon evolutionär prediktion. Sanningen är att den totalt dominerande synen hela tiden har varit att det mesta av DNA:t är skräp eftersom det betraktats som nödvändigt för att evolutionen skulle vara möjlig.
Det som skedde 2012 kom därför som en total chock för det evolutionära etablissemanget.ENCODE
Låt oss därför titta närmare på E.s andra argument, det om ENCODE-projektet. 2012 presenterade ENCODE-projektet sin första rapport som utgjorde slutsatserna från fler än fyra hundra forskare från dussintals universitet, forskningsinstitut och laboratorier från USA, Europa och Asien. Det tidigare omnämnda HUGO-projektet hade fokus på den del av människans genom som kodade för proteiner. Det nya projektet satte i stället strålkastarljuset på återstoden av arvsmassan, ”skräpet”. Slutsatserna: ungefär 80 procent av genomet visade tecken på funktion. Med tanke på att man bara studerat en blygsam andel av alla de olika celltyper som kroppen består av menade en talesperson att den andelen skulle kunna närma sig hundra procent.
E. använder fyra sidor (96-99) till att beskriva projektet i mycket kritiska ordalag. Det är inte helt oväntat. De flesta evolutionsbiologer reagerade på tre olika sätt när ENCODE-resultaten publicerades. En del blev arga, andra förnekade data, ytterligare andra försökte skriva om historieböckerna. E. kombinerar alla tre. Men innan jag beskriver E.s kritik ska jag ge en bakgrund till vad det var som ENCODE visade.
Den läsare som glömt sin naturkunskap från gymnasietiden får här en kort uppdaterad repetition.60
De gener (delar av DNA) som kodar för proteiner översätts med hjälp av en grupp molekylära maskiner som kallas transkriptaser till en annan informationsbärande molekyl som kallas RNA, med en delvis annan kemisk sammansättning. Dessa så kallade budbärar-RNA-molekyler (mRNA) lämnar cellkärnan. Ute i cytoplasman redigeras mRNA-molekylerna på olika sätt så att det av en enda ursprunglig mRNA-molekyl kan bildas tusentals olika varianter genom en sinnrik klippa-klistra-procedur som möjliggörs av den kanske mest komplexa molekylära maskin som existerar, den så kallade spliceosomen.61 Dessa redigerade mRNA-varianter utgör recept på de enskilda proteinerna som cellen behöver vid ett visst givet tillfälle. En annan molekylär maskin – ribosomen – fäster med sina två underenheter på en mRNA-sträng och glider utmed den, medan den läser av nukleotidsekvensen (”RNA-bokstäverna”) och tillverkar en polypeptid (proteinmolekyl) genom att fästa aminosyror vid varandra i den ordning som mRNA-sekvensen (”receptet”) anger. Fler detaljer får du via internet.
De proteiner som bildas motsvarar de olika bitarna i en Lego®-förpackning. Tänk dig att du för en förmögenhet köper in några kilo legobitar och lägger dem i en stor hög på golvet. Av dessa legobitar i olika storlek, form och färg kan du nu börja bygga allehanda saker: bilar, båtar, flygplan, name-it. För att göra det behöver du kreativitet och fantasi. Eller instruktioner. De senare medföljer varje paket, och kanske brukar du slänga dem i papperskorgen i tron att de inte är till någon nytta. Man brukar ångra sig.
Observera att man av i stort sett samma legobitar kan bygga en mängd olika konstruktioner. Det är precis på samma sätt i den levande världen. Av i stort sett samma proteiner, kodade av i stort sett samma sekvenser (gener) i DNA bygger ”naturen” människor, schimpanser, möss och allt annat levande. Det finns någonstans mellan 250-500 gener som kodar för proteiner som förekommer i samtliga jordens organismer (så långt man kartlagt dem), oavsett om vi talar om bakterier, gurkor, makrillar eller människor.
Men, undrar du kanske, om alla olika varelser är uppbyggda av i stort sett samma proteinmolekyler – vad är det då som gör att de monteras ihop på så helt olika sätt? Svaret kan ju inte ligga i det DNA som är proteinkodande. Nej, det måste ligga någon annanstans. Vad finns det att välja på? Svaret är givet – det måste naturligtvis ligga i det DNA som inte är proteinkodande. Skräp-DNA! Det är där hemligheten döljer sig som skiljer oss från schimpanser, eller för all del myggor. I det icke-proteinkodande DNA:t finns nämligen instruktionerna för hur varje särskild levande varelse ska monteras. Vi talar om de genetiska instruktionsböckerna, de som evolutionsbiologerna sedan 40 år har kastat i papperskorgen som skräp!
ENCODE-projektet studerar alltså instruktionsböckerna. E. skriver kapitlet i förväntan att dessa inte ska vara så omfattande som det tycks och att ENCODE-forskarna har överdrivit sina resultat.
Hur går det till när det icke-proteinkodande DNA:t ser till att en människa blir just en människa och ingenting annat? Det sker tack vare att det i stället för proteiner framför allt kodar för RNA-molekyler som inte utgör recept på proteiner utan i stället har reglerande funktioner. Dessutom döljer sig där en rad andra nödvändiga molekyler som transposoner och repetitiva sekvenser. Tillsammans möjliggör de att ett ägg av en enda cell kan bli en människa bestående av hundratals olika sorters celler. Den där instruktionsboken, helt enkelt.
Nu till E.s kritik mot ENCODE. Den går ut på följande. (Beklagar den här djupdykningen med tanke på dig som läser och inte är naturvetare):
1. att man konstaterar att en viss gen är aktiv behöver inte innebära att den har någon funktion. (s 97-98)
2. att en sekvens i DNA översätts till RNA (som miRNA) inte heller behöver innebära funktion, för det är så få RNA-molekyler att det ibland är färre än en per cell. (s 97)
3. Att helt slumpartade DNA-sekvenser också kan ge sken av funktion. Alltså är resultaten överskattade. (s 98)
Mina kommentarer:
1. Alla aktivitet innebär kostnader i from av energi och material och därmed en negativ fitness (fortplantningsnackdel). Selektionen borde över tid ha eliminerat sådana källor till resursslöseri.
2. Att RNA ofta förekommer i så låga koncentrationer som ”mindre än en molekyl per cell” beror på att det handlar om genomsnitt för hela organismen. Kroppen består av 30 biljoner celler så antalet molekyler i en specifik celltyp62 i en vävnad kan ändå vara betydande. Dessutom kan det faktiskt räcka med enstaka molekyler av signalsubstanser för att påverka genregulatoriska nätverk.63, 64
3. Med tanke på att ingen räknar med en 100-procentig perfektion i nutida DNA, så kan det absolut finnas ett visst ”brus” i form av produktion av onödigt RNA, men det vore att kasta ut barnet med badvattnet att dra alltför långtgående slutsatser av det.
Enligt E. är slutsatsen från ENCODE-projektet att en stor andel av vårt DNA fortsatt tycks vara icke-funktionellt. (s 99)
Notera kontrasten mot vad molekylärbiologerna Mattick och Amaral skriver i sin bok RNA, the Epicenter of Genetic Information (2022): 65
Även om historien fortfarande utvecklas, drar vi slutsatsen att genomet hos människor och andra komplexa organismer inte är fullt av skräp, utan snarare är mycket kompakta informationspaket som till stor del ägnas åt specifikationen av regulatoriska RNA. Dessa RNA styr differentierings- och utvecklingsförloppet, ligger till grund för hjärnans funktion och förmedlar transgenerationellt minne av erfarenheter, vilket till stor del strider mot långvariga uppfattningar om genetisk programmering och dogmerna i evolutionsteorin.
Notera speciellt den sista meningen: ”Slutsatsen … strider mot långvariga uppfattningar om … dogmerna i evolutionsteorin.”
Man kan tycka att evolutionister borde ha lärt sig läxan från 1900-talets lista av spårlöst försvunna rudimentära organ. Det var ingen brist på funktion, det var brist på kunskap. Historien håller på att upprepa sig inför våra ögon. Något enstaka undantag, som till exempel visdomständer hos oss med nutida födovanor bekräftar regeln. Det finns ingen anledning att inte samma sak skulle gälla på DNA-området. E. argumenterar i hopp om att behålla föreställningen om ”skräp-DNA", men det lär vara förgäves. Det går nämligen aldrig att bevisa icke-funktion, bara dess motsats.
Till sist, E.s tredje och sista argument, att det egentligen kvittar om det visar sig vara litet eller mycket skräp i vårt DNA.
Om E. verkligen tyckt att det hade kvittat så hade han rimligtvis inte lagt ner all den här mödan på att utveckla sin kritik. Men han inser samtidigt att det dagligen strömmar in rapporter om nya funktioner i ”skräpet” och att risken att bli inmålad i ett hörn är överhängande. Det behövs en nödutgång. E. löser knuten genom att referera till en källa från 2020 (E.s not 49 s 96) där man lanserar hypotesen att hela genomet skulle kunna vara funktionellt eftersom mutationernas effekt inte är så stor som man tidigare trott.
Han är väl medveten om evolutionsteorins behov av ett skräp-DNA-upplag för att begränsa mutationers negativa effekter och som en tummelplats för uppkomsten av nya anlag. Men skulle det utrymmet försvinna så återstår bara att oskadligförklara mutationerna. Han är ju trots allt redan inne på den linjen så det skulle ändå inte förändra så mycket i sak.
Håller du med E. om att skräp-DNA-metaforen är relevant? Det gör inte jag.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 15. Fjärde argumentet: Komplexitetsargumentet
Vad menar vi när vi säger att någonting är komplext? Till vardags menar man som regel att någonting är väldigt invecklat och att det är svårt att få rätsida på det, som trassel på fiskelinan när man är ute och dörjar makrill. Det där är bara en aspekt på saken och en som inte är i fokus för designteoretiker som William Dembski, Stephen Meyer och Michael Behe. Anledningen till det är att komplexa saker händer hela tiden av helt naturliga anledningar. Som det där lintrasslet. Eller den kemiska sammansättningen på de organiska ämnena i en meteorit. Eller ordningen på talen när man kastar en tärning upprepade gånger. Slumpen och naturlagarna är fullt tillräckliga för att förklara komplexa eller osannolika saker som uppstår.
Inom ID talar man om två andra komplexitetsbegrepp. Dels specificerad komplexitet (Dembski och Meyer) och dels oreducerbar (E. översätter det irreducerbar) komplexitet (Behe). Vi återkommer till begreppet specificerad komplexitet under argumentet Förklaringsfiltret längre fram (kap 18). Här och nu är det Michael Behes oreducerbara komplexitet som E. har invändningar mot.
Oreducerbar komplexitet
I sin bok Darwin’s Black Box från 1996 diskuterar biologen Michael Behe funktionella system. Han illustrerade det med den numera klassiska musfällan. En musfälla består av ett begränsat antal komponenter (delar). De kan placeras på lite olika sätt och antalet komponenter kan variera inom vissa snäva gränser, men det finns en gräns för hur mycket konstruktionen kan reduceras (minskas ned) innan dess förmåga att fånga möss upphör. Den enklaste fungerande versionen av musfällan är enligt Behe oreducerbar. Eftersom funktionen att fånga möss uppstår först när systemet med dess olika komponenter finns på plats så kan den, menar han, inte ha uppkommit genom en stegvis selektionsprocess där man hela tiden valt ut den musfälla som var bäst på att fånga möss, eftersom det inte existerade någon musfångande funktion att välja ut.
Behes poäng var att om motsvarande strukturer går att påvisa i naturen, så skulle inte heller dessa kunna ha uppstått genom någon stegvis selektionsprocess (läs: evolution). Gör man det? Tveklöst, och Behe ger själv ett antal exempel i sin bok. Som tidigare nämndes i samband med design-detektering (s 30) föredrar Behe att definiera specificerad komplexitet som ”ett ändamålsenligt arrangemang av delar”.
Vi har ägnat magasinet Genesis ett helt nummer åt system. Det rekommenderas varmt för den som vill få en djupare förståelse för Behes argument.66
Tveksam systemkritik
System är kännetecknande för alla jordens livsformer. System som är integrerade i andra system som i sin tur är integrerade i ytterligare andra system.67 Sådana arrangemang hos levande varelser illustrerar hur osannolikt (läs: uteslutet) det är att evolutionära processer skulle kunna förklara deras uppkomst. Vi inser alla omedelbart hur kraftfullt det här argumentet är.
Hur tar sig nu E. an det här evolutionära dilemmat? Han ger två argument för att det inte finns någon substans i komplexitetsargumentet:
1. Systemet får inte ha förändrats över tid så att det till exempel hade fler komponenter förr i tiden.
Men att strukturer nedmonteras har aldrig varit problemet. Det sker ständigt i naturen och är – som vi såg när vi behandlade mutationer (kap 8) – den vanligaste orsaken till fördelaktiga mutationer, vilket är ett stort evolutionärt dilemma. Som vi tidigare konstaterat – evolutionen behöver inte förklara avveckling mot lägre komplexitet utan utveckling mot högre. Ett sådant system med fler komponenter som E. refererar till är heller inte oreducerbart, för i så fall hade det inte gått att reducera med bibehållen funktion. Ifall det såg annorlunda ut i det förgångna är alltså irrelevant för frågan.
2. Varje komponent i systemet får inte växla till en annan funktion utan bara ha en enda oföränderlig sådan.
Men mig veterligen finns det inte en enda observation i den vetenskapliga litteraturen av att en funktion i ett levande system setts ”växla till någon annan”. Däremot finns det några få exempel på evolutionära berättelser om sådant,68 men det är naturligtvis en helt annan sak. E.s slutsats är:
[Ä]ven om vi idag finner att ett system skulle vara icke-reducerbart komplext i Behes mening, finns det i princip ingenting som hindrar att det kan ha evolverat via bekanta evolutionsmekanismer. (s 101)
Men genom att (1) förlora komponenter och (2) utan att ha någon observation till stöd för sin uppfattning så är E.:s slutsats helt och hållet baserat på en blind tro på vad mutationer och naturligt urval kan åstadkomma. Och vi har redan sett att de båda mekanismerna bevisligen är djupt destruktiva till sin natur.
E. menar att det är Behe som måste lägga fram evidens för sin sak och inte evolutionsbiologerna som ska behöva bevisa att Behe har fel:
I vetenskapliga sammanhang är det den som gör ett påstående som också bär bevisbördan. (s 102)
E. har naturligtvis rätt i att den som hävdar att något är omöjligt normalt sett bär bevisbördan. Och det stämmer också att det kan vara orimligt att kräva detaljbeskrivningar av evolutionära förlopp.
Däremot är det fullt möjligt – och fullt vetenskapligt – att hävda att en process är osannolik eller begränsad utan att man för den skull behöver känna till alla detaljer. Man måste till exempel inte undersöka alla föreslagna evighetsmaskiner för att kunna påstå att de inte fungerar. Behe behöver inte heller visa att alla evolutionära processer är orimliga. Det han har gjort är att presentera generella argument för att mutationer och selektion (med avseende på en ännu icke existerande egenskap) inte är tillräckliga som förklaring till uppkomsten av vissa strukturer i naturen.
Vi har en omfattande erfarenhet av att det bara är intelligenta aktörer som skapar system och att naturliga processer inte förmår göra det. Det är därför högst rimligt att en del av bevisbördan vilar på den som påstår motsatsen.
Så är Behes påstående verkligen ovetenskapligt? Är det mer vetenskapligt att hävda att något har hänt än att något inte har hänt? I så fall varför?69
Som lite kuriosa vill jag påminna om Charles Darwin som 1859 skrev i sin berömda bok Om arternas Uppkomst:
Om det kunde bevisas att det fanns något komplext organ som omöjligen kunnat bildas genom ett stort antal, på varandra följande, små förändringar, skulle min teori helt och hållet bryta samman. Men jag kan inte hitta något sådant fall.
E. återger också det här citatet på s 60. Vi kan konstatera att Darwin lade över bevisbördan på sina kritiker trots att det var han själv som gjorde ett vid den tiden väldigt kontroversiellt påstående. Ska man döma efter E.s mått var detta ett bedrägligt förfarande av Darwin.
E. ägnar sedan ett par sidor åt att kritisera argumenten i Behes andra bok, The Edge of Evolution (2007). (s 103-104) I den här boken vill Behe pröva evolutionens förmåga att genom mutationer och naturligt urval testa sig fram för att hitta lösningar på problem. Som alla evolutionära processer förutsätts det här naturligtvis vara en blind och planlös process eftersom mutationer anses ske helt slumpmässigt.
Ju fler individer en population av en viss organism består av, desto större är chanserna för att en viss fördelaktig mutation ska råka uppstå. Det betyder förstås att om man jämför en population av elefanter eller en population av mikroorganismer som delar samma gen (vilket inte är så ovanligt som man föreställer sig), så måste man vänta orimligt länge för att hitta den bland elefanterna på grund av deras långa generationstid och fåtaliga ungar.
Malaria
Behe valde att studera data om Plasmodium falciparum, den blodparasit som förorsakar malaria. Den förekommer hos ungefär en miljard (109) människor och i ett ungefärligt individantal per person på ungefär en biljon (1012). Det ger en global population på 1021. I en så stor population uppkommer i stort sett alla tänkbara mutationer i varje generation.
Det finns flera olika malariamediciner, en del effektivare än andra. Problemet är att parasiterna kan utveckla resistens som gör att mediciner blir verkningslösa. En av medicinerna heter atovakvon, en annan klorokin. Resistens mot atovakvon är hundra miljoner gånger vanligare än mot klorokin. Den uppträder i ungefär var tredje person som är bärare av malaria, medan man räknar med att resistens mot klorokin bara verkar ha uppkommit mindre än tio gånger de senaste femtio åren.70 Behe ville förstå varför. För att en malariaparasit ska utveckla resistens mot atovakvon räcker det med en (1) specifik mutation i ett visst protein. Men för att en parasit ska bli resistent mot klorokin krävs det två (2) specifika mutationer. Behe konstaterade att det är detta som är hemligheten bakom den extrema sällsyntheten med klorokinresistens. Sannolikheten att få en specifik mutation på ett visst ställe i en viss gen är förhållandevis överkomlig, men om kravet för resistens är två specifika mutationer på bestämda ställen så minskar sannolikheten drastiskt. Det beror på att sannolikheter för oberoende händelser alltid multipliceras.71
Vad spelar det här för roll undrar du kanske? Jo, för små populationer som till exempel människor, så finns det inte på långt när så många chanser till lyckträffar för mutationer som i populationen av malariaparasiter ens under miljarder år. Märk väl att Behes resonemang inte handlar om spekulationer, utan baserar sig på väldokumenterade fakta om kända mutationsfrekvenser och kända hälsodata. På den grunden har han visat att det skulle vara omöjligt att bygga upp genomen hos jordens levande varelser eftersom det kräver enorma mängder av lyckosamma mutationskombinationer. Evolutionsteorins förklaringskraft är alltså totalt försumbar i relation till hur världen de facto ser ut.
Kritik mot The Edge of Evolution
Det här var bakgrunden. Hur angriper då E. den här utmaningen? Det blir för mycket citerande om jag ska återge allt, så jag väljer några representativa formuleringar:
Mutationerna måste ha skett samtidigt, säger han [Behe], eftersom en ensam mutation högst sannolikt hade varit negativ och skulle rensats bort av den naturliga selektionen. (s 104)Om nu mutationer kan ske i sekvens i stället för samtidigt ökar det dramatiskt sannolikheten för att en mutation kan följas av ytterligare en. (s 104)
Diskussionen avgjordes 2014 [Summers et al] … Det gav kritikerna rätt. Arbetet visade att det finns ett flertal olika mutationer som kan bidra till resistensutveckling och också flera vägar för hur resistens kan utvecklas. (s 104)
De flesta mutationerna var positiva (tio stycken) eller neutrala (åtta). Ett mindre antal (fem) var svagt negativa. Det var inte alls så att positiva och neutrala mutationer var helt osannolika, som Behe tänkte sig. (s 105)
Därför behövdes inga osannolika dubbelmutationer. (s 105)
E.s argument är ett eko av den kritik som några andra kritiker riktar mot Behes bok. Även de menar att klorokinresistens hos malariaparasiten inte alls behöver uppkomma genom ett antal mutationer på en och samma gång, utan i stället genom en serie av mutationer som steg för steg ger parasiten lite högre grad av resistens. (första, andra och femte citatet ovan). Alltså ungefär som att man i stället för att behöva hoppa åtta meter meter ut till en holme, så kan man hoppa via ett antal stenar en kortare bit i taget.
Michael Behes eget svar på kritiken
Behe svarar på den kritiken med hjälp av följande analogi, fritt återgiven:
Du ska delta i ett lotteri där det handlar om att gissa en följd av sju tal. De sista fem talen kan vara vilket heltal som helst mellan noll och tre, och i vilken inbördes ordning som helst. De två första talen kan vara vilket heltal som helst mellan noll och en miljard.
Vilka av de sju talen är det som är största hindret för dina chanser att vinna?
Givetvis de två första.
Det vetenskapliga arbetet från 2014 som E. menar ”avgör diskussionen” visar att det är just detta som är problemet med uppkomsten av klorokinresistens: För att parasiten ska kunna utveckla någon resistens över huvud taget, så krävs det två specifika mutationer. När det osannolika väl har inträffat (de två första talen i talföljden blir rätt) – först då kan det börja ske en selektion av ytterligare mutationer som ökar graden av resistens ett litet steg i taget (vilket motsvarar de fem sista siffrorna i talföljden, de som var ganska lätta att få till).
Huruvida det krävs två eller tio mutationer, ifall de är positiva, neutrala eller negativa och om det finns en eller tusen mutationsvägar att nå fram till resistens hos malariaparasiten är helt oväsentligt i sammanhanget.
Varför då?
Jo, vi har nämligen facit i hand baserat på övertygande statistisk evidens: oavsett hur resistensen mot klorokin uppstår så uppkommer den bevisligen i storleksordningen 1 på 1020 (plus minus ett par tiopotenser).
Om det är lite svårt att greppa så tänk dig följande fiktiva scenario: Det är orienteringstävling med 10 000 deltagande som alla ger sig väg ut i skogen. De saknar karta och kompass utan får lita på sin intuition. Det är mulet så de har ingen sol att gå efter och de har inte gått i scoutverksamhet så de har ingen aning om att man kan använda myrstackar till att bestämma väderstrecken. Deras sökning är helt slumpmässig, precis som mutationer. En mil bort finns målet. Arrangörerna spekulerar i hur stor sannolikheten är att en orienterare hittar till målet. Man gör mängder av beräkningar som alla varierar beroende på vilka antaganden man gör om hur många stigar som leder fram till målet (hur många vägar till resistensutveckling), hur snabbt man springer och i vilka riktningar (positiva, negativa eller neutrala mutationer). Arrangörerna undrar om de någonsin kommer att vara möjligt att räkna ut den där sannolikheten att nå målet.
Det är då någon kommer på den geniala idén.
Vi kollar hur många som kommit fram till målet!
Det var 7.
7 av 10 000. Det är exakt 0,07 procent!
Det är samma sak i vårt fall. Vi har facit från medicinsk statistik. Chansen är ungefär 1 på 1020!
Men det finns mer att säga. Den källa som E. refererar till, Summers (2014) bekräftade Behes slutsats att det krävs minst två specifika mutationer för att erhålla resistensen. 72 Den ena (i position 76) var exakt den som Behe förutsade.73 Det har visat sig att den inte är neutral som E. påstår, utan skadlig.74 Det är just därför som en annan mutation (x eller y enligt Summers) måste till samtidigt. Och det är denna omständighet som gör dubbelmutationen så sällsynt förekommande i populationen av Plasmodium som en på 1020. Om den varit neutral skulle dubbelmutationen ha varit avsevärt vanligare än den faktiskt är med facit i hand!
Behe har därmed bevisat att de där två specifika första mutationerna är en grundförutsättning för att de ska kunna uppstå en svag resistens hos Plasmodium. När denna väl har uppkommit är det inget som helst problem för det naturliga urvalet att selektera fram ett antal mutanter som har ännu bättre resistensegenskaper.75 Det är först nu det finns ett antal olika möjligheter att nå den fulla resistensen. Men, som sagt, utan de där båda mutationerna – ingen resistens alls. Det är därför som klorokinresistensen är så extremt sällsynt. Behe har rätt. E. fel.
Det kvittar alltså helt och hållet hur många av mutationerna som är positiva, negativa eller neutrala eller om det finns en eller tusentals olika vägar för att nå målet.
Vad innebär det här för E.s resonemang? Svaret är: Resonemanget är totalt irrelevant för frågan.
De båda andra exemplen
Nästa exempel Erkell levererar är från en grupp forskare vid Uppsala universitet (s 105), som odlat Salmonella-bakterier som försetts med ett bifunktionellt enzym. Det innebär att det har förmågan att tillverka två olika aminosyror med låg effektivitet. Under loppet av ett år kunde man isolera stammar som muterat så att de var effektivare på att tillverka antingen det ena, det andra eller båda två aminosyrorna. Det här är helt i enlighet med de fem sista talen i lotteriraden ovan. Mutationer och naturligt urval är som sagt alldeles utmärkta på att finjustera saker som enzym. Men under en enda förutsättning: att det från början finns en aktivitet. Utan den får man ingen ”evolution”.
E.s sista exempel handlar om bakterieätande virus, så kallade fager, i det här fallet lambda-fager (s 105-106). Av de fyra mutationerna så innebar de tre första en ökad bindningsstyrka till cellyteproteinet LamB. Den fjärde gjorde också det, men kunde dessutom binda till proteinet OmpF. Eftersom både LamB och OmpF har samma 3D-struktur är det inte särskilt konstigt att virusproteinet kan binda båda. Det är visserligen frågan om en för viruset fördelaktig mutation, men till sin karaktär är den inte nyskapande utan en modifiering av en redan befintlig funktion.
Det visar sig att värdcellerna lyckades lura lambda-fagerna genom att mutera även OmpF-proteinet. En mutation stängde av genen för produktion av proteinet. Konsekvensen blev att bakterien inte längre kunde släppa in sockerarten mannos och använda den i sin ämnesomsättning. Med Behes egna ord:
Så i slutändan återstod den muterade bakteriofagen lambda, som fortfarande var oförmögen att invadera de flesta E. coli-celler, samt muterad E. coli, nu med trasiga gener som tog bort dess förmåga att metabolisera maltos76 och mannos. Det verkar som om den darwinistiska evolutionen tog ett litet steg åt sidan och två stora steg bakåt.”77
E. konstaterar avslutningsvis:
Konsekvensen är att det aldrig går att veta att en förmodat irreducerbart komplex struktur verkligen är irreducerbar, så begreppet är inte meningsfullt. (s 106)
Men det stämmer alltså inte. Just de vetenskapliga observationer som E. refererar till understryker att evolutionen verkligen har gränser. Vi har malariaparasiten att tacka för det. Det här är naturligtvis inte att argument från okunnigheten, som han påstår. Tvärt om är det en vetenskaplig slutsats baserad på beräkningar utifrån faktiska observationer.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 16. Det femte argumentet: Det kombinatoriska argumentet
E. inleder sin behandling av det här argumentet med att presentera läsarna en nidbild (halmdocka) av det (s 108). När han sedan har lagt fram sina argument avrundar han kapitlet med att avfärda det som – just det – en halmdocka (s 111).
Låt mig först förklara bakgrunden för dig som inte läst boken. För dig som läst den blir det en repetition:
Proteiner består av långa kedjor av aminosyror. Av dessa finns det 20 olika sorter,78 som skiljer sig från varandra genom olika så kallade sidokedjor med olika kemiska och fysikaliska egenskaper. Aminosyrorna fungerar ungefär som bokstäverna i vårt alfabet. Olika kombinationer av bokstäver ger olika slags litteratur, och motsvarande ordning på aminosyrorna ger proteiner med olika funktioner, som rörelse (muskelprotein) byggmaterial (t ex kollagen och keratin) eller för att reglera kroppens ämnesomsättning (som enzymer).
De där sidokedjorna på aminosyrorna gör att olika delar av ett protein får olika egenskaper – vissa delar attraherar vatten, andra attraherar i stället fett, vissa attraherar positiva elektriska laddningar andra negativa, vissa är sura, andra basiska och så vidare. Allt det här gör att ordningen på aminosyrorna påverkar möjligheten för en viss del av aminosyrakedjan att attrahera en annan del av sig själv. En kedja av aminosyror som är helt slumpmässig i sin ordningsföljd (sekvens) kommer med största sannolikhet att bete sig som en överkokt spagetti i en kastrull med kokande vatten. Den är lös och sladdrig. Sådana aminosyrakedjor – om de mot förmodan skulle bildas i en cell, vilket vore slöseri med resurser – skulle snabbt sönderdelas (på kemispråk hydrolyseras) tillbaka till fria aminosyror. Det finns en naturlag som ombesörjer det – massverkans lag.
Men vissa kombinationer av aminosyror möjliggör för den där spaghettitråden att attrahera sig själv på ett sådant sätt att den bildar områden i form av antingen en spiral eller en flat yta. Det kallas för dess sekundärstruktur. Det gör aminosyrakedjan stabil, så att den inte genast sönderdelas till sina beståndsdelar.
Eftersom även de delar av en aminosyrakedja som inte bildar spiraler eller ytor attraherar varandra så kommer den dessutom att bilda en ”klump” med en viss tredimensionell form (tertiärstruktur). Det förutsätter förstås att ordningen på aminosyraenheterna där medger det. Det är den här formen på klumpen som ger molekylen möjlighet att ha en viss funktion i cellen. Det är först då som aminosyrakedjan får lov att kallas för ett protein.79
Utan att diskutera procentandelar kan vi konstatera att det inte är alla kombinationer av aminosyror som leder till att det uppstår ett funktionellt protein.
Det låter här som att det vore enkelt att få aminosyror att slå ihop sig till kedjor, men så är det definitivt inte, i synnerhet inte på ett sätt som ger dem förutsättningar att utöva en funktion. Det här beror på det jag skrev nyss, att det finns en kemisk lag som tenderar att spontant sönderdela aminosyrakedjor i vattenmiljö i stället för tvärt om.80 Och det finns ytterligare en anledning – i praktiken ett monumentalt problem för evolutionsteorin som E. inte nämner ett enda ord om. Jag kommer att behandla det i slutet av det här avsnittet.
När man väl har ett funktionellt protein, så har man visat experimentellt att det som regel bara krävs 3-15 utbyten av aminosyror (mutationer) för att dess funktion helt ska upphöra. Det vanligaste är en enskild mutation försämrar funktionen, men det förekommer även att en mutation kan förbättra funktionen, speciellt om miljön förändras. Det är just det senare som möjliggör för det naturliga urvalet att göra individer bättre anpassade vid miljöförändringar.
Det här var en lång introduktion, men det är viktigt att ha bakgrunden klar för sig.
Det missuppfattade argumentet
När E. introducerar det kombinatoriska argumentet så börjar han med att berätta hur osannolikt det är att slumpmässigt kombinera ihop aminosyrorna i en kedja med 300 aminosyror på ett visst sätt (s 108). Det motsvarar att kasta 300 tjugosidiga tärningar så att ordningen på siffrorna som uppstår (det förutsätts att de hamnar i en lång rad) stämmer överens med en viss i förhand bestämd ordningsföljd. Det skulle motsvara att slumpa fram ett visst specifikt protein med en viss specifik funktion, som till exempel enzymer. Att detta är astronomiskt osannolikt är förstås bara förnamnet. Det här menar E. utgör ID:s argument för att proteiner är designade. Och, som han senare skriver:
Ingen biolog vid sina sinnens fulla bruk tror dock att komplexa proteiner bildas i ett slag genom att evolutionen blint letar efter nya fungerande proteiner i denna ofattbara mängd av möjliga kombinationer. (s 108)
Lite indirekt alltså: Personer som tror det är inte vid sina sinnens fulla bruk. Följaktligen är ID-anhängare hiskeligt korkade.
Med det här kapitlet vill docent E. få läsaren att inse att nutida proteiner i stället har uppkommit genom en steg-för-steg-process. Det gick till så här, menar han: (s 109) Det började med korta RNA-strängar på sådär fem nukleotider eller rentav enskilda aminosyramolekyler med svag enzymfunktion. Nu startade en spontan urvalsprocess som successivt ledde till allt större enzymmolekyler, så att det till slut resulterade i de nutida som ofta har tusentals aminosyror i sina kedjor.81
En novell
Innan vi går vidare vill jag göra en analogi. E. avskyr analogier (s 34). Du kommer att förstå varför:
En novell är en kort text som i omfattning kan jämföras med ett protein i en levande organism, till exempel ett enzym som ser till att DNA-molekylen kopieras på ett korrekt sätt och korrekturläser resultatet så att det inte uppstår för många fel (mutationer) i förhållande till originalet. Ett sådant enzym måste ha existerat från allra första början, eftersom den förhållandevis blygsamma82 mängd genetiska information som fanns där inte skulle försvinna efter några få celldelningar.
Hur stor är chansen att någon sorts slumpmässig ”händelse” vid ett tangentbord skulle råka åstadkomma den där novellen inom loppet av några sekunder? Praktiskt taget noll, eller hur? Där har E. alltså helt rätt. Han fortsätter nu sitt resonemang:
Det är inte så en novell uppstår. I stället byggs den upp stegvis genom att bokstav fogas till bokstav. Den behöver alltså inte alls uppstå i ett enda nu. Hemligheten ligger i att tillkomstprocessen sker i små steg efter varandra under långa tidsrymder. Alltså – det behövs ingen mystisk författare!
Känns det som en rimlig slutsats?
Det är i och för sig helt riktigt att en novell uppkommer stegvis genom att bokstäverna placeras där från vänster till höger. Men – och det är inget litet men – utesluter det verkligen behovet av en författare? Det är nämligen vad E. påstår. Självklart inte.
Det är däremot sant att novellen uppstår genom selektion. Varje bokstav väljs ut bland 29 möjliga. Men det är en medveten, intelligent, personlig, levande varelse som gör urvalet,83 inte blind slump.
Det som möjliggör novellens existens är alltså inte en planlös, stegvis process i sig – vilket E. vill låta påskina – utan den intelligent guidade skrivprocess som ständigt har det framtida målet för ögonen. Utan både mål och förutseende blir processen ett planlöst irrande som i stort sett inte kommer att leda i någon riktning alls, och chansen att nå målet blir helt och hållet beroende av slumpen. Det är då som de där oddsen blir så astronomiskt små att nå målet. Selektion måste alltid ha en riktning när det gäller att kunna leda till någonting.
Dawkins analogi
Det är inte bara jag som tar till analogier för att förklara saker i den levande världen. I sin ovan nämnda bok Den blinde urmakaren ville Richard Dawkins demonstrera hur kraftfullt det naturliga urvalet är när det gäller att skapa ny information steg för steg.
För att göra det hade han skapat ett datorprogram med förmåga att skapa en kort strof ur Shakespeares Hamlet, närmare bestämt ”METHINKS IT IS LIKE A WEASEL”. Hans program gjorde det genom att slumpmässigt generera bokstäver (som motsvarar mutationer) och sedan selektera dessa på grundval om de råkade bli "rätt" eller inte (”naturligt urval").
Hans datorsimulering ledde efter 43 "generationer" fram till sitt mål – den aktuella Shakespearefrasen.
Men att hans exempel inte har någon bäring på frågan om den biologiska informationens uppkomst behöver väl knappast påpekas. Det enda hans intelligent designade dator åstadkom var att återskapa den information som redan från början var placerad där som mall (målsekvensen)! 84
Dawkins exempel är därför fullständigt irrelevant som parallell till evolutionen i termer av mutationer och selektion. Däremot fullt relevanta för att illustrera en skapelseakt! Dawkins datorprogram utgör en metod för att skapa en slutprodukt som redan från början finns i konstruktörens sinne. Det är ju faktiskt precis så både jag och E. gör när vi författar våra artiklar och böcker. Vi väljer bland en uppsjö av ord, och när vi finner det ord som stämmer bäst med vad vi vill uttrycka så ser vi till att snabbt få det på pränt och spara ner den på hårddisken. "Slumpen" (hjärnstammen?) genererar orden och vi själva selekterar fram de som är mest "adaptiva".
Naturligtvis ligger det ett visst mått av sanning i det E. skriver, nämligen att ID-företrädare anser att fungerande proteiner är på tok för specifika till sin sammansättning för att kunna ha uppkommit genom mutationer och naturligt urval, men det är just det som är det förrädiska med missrepresentationer – den där blandningen av sanning och osanning. ID hävdar inte att ett protein skulle poppa fram i ett endaste nu, och det är därför varken någon halmdocka eller någonting man angriper utifrån okunnighet om hur världen fungerar. Det är i stället någonting med massivt stöd i evidensen. Låt mig nu bevisa det.
Först måste vi klargöra vad det här så kallade kombinatoriska ID-argumentet egentligen handlar om. Vi börjar med en språkfråga (DNA-sekvenser är ju faktiskt en sorts text med en egen grammatik och som kodar för proteinernas sammansättning och egenskaper):
Man kan undra hur många bokstavskombinationer som ger en begriplig mening i förhållande till de som inte ger det. Jag vet ingen sådan undersökning för svenska språket, men det har gjorts för engelskan. Det visar sig att det för varje meningsfull engelskmening på tolv bokstäver finns 1014 (hundra tusen miljarder) meningslösa bokstavskombinationer. Det här förhållandet ökar inte linjärt utan exponentiellt i takt med att meningarna blir längre, så för varje engelsk mening som är 100 bokstäver lång finns det 10100 nonsensmeningar.85
På liknande sätt kan man fråga sig hur många kombinationer av aminosyror som ger ett fungerande protein i förhållande till hur många som leder till lika långa men funktionslösa proteiner. Det är antagligen omöjligt att få reda på det. Men det går faktiskt att på experimentell väg göra en konservativ uppskattning om saken, alltså säkert visar på en högre andel än den verkliga.
En proteinforskare vid Cambridge i England vid namn Douglas Axe tog sig an det problemet genom att studera proteiners stabilitet. Stabilitet hos en kedja av aminosyror är inte synonymt med att den har en funktion (det är först då den får kallas protein), men det är en grundförutsättning för funktion.86 Det här studerade Axe och hans forskningsteam med hjälp av en metod som går under namnet ”riktad mutagenes”. Den innebär att man skapar väldiga mängder med snarlika proteinkopior där man bytt ut aminosyrorna i vissa delar av proteinet och ser hur det påverkar stabiliteten. Studien visar att om man utgår från ett stabilt protein, så krävs det bara någon eller några få mutationer för att stabiliteten ska påverkas negativt eller helt upphöra. Som regel förlorar ett protein sin stabilitet fullständigt efter 3-15 mutationer. Axe och hans team kunde efter fjorton års studier (1990-2003) dra slutsatsen87 att det för varje stabil sekvens med 150 aminosyror finns 1077 aminosyrasekvenser som är instabila och som därför saknar förutsättningar att ha någon som helst biologisk funktion. Det motsvarar antalet möjliga sifferkombinationer till ett kodlås med 77 sifferhjul (det brukar ju vara tre till sex stycken på ett cykellås. Det låter nästan hanterbart. Men skenet bedrar – i realiteten är det ett tal som är hundra miljoner gånger fler än det finns atomer i vår galax, Vintergatan.
Och som sagt, det här är en kraftig underdrift. Vi vet inte hur stor andel av stabila strukturer som dessutom har en funktion. Till råga på allt handlar det här om ett mycket kort protein. I medeltal är de dubbelt så långa, och precis som i bokstavsfallet så ökar osannolikheten exponentiellt med antalet.
Så av 1077 relativt korta aminosyrakedjor är det bara en enda som inte beter sig som en överkokt spaghetti i den där kastrullen med kokande vatten. I en naturlig miljö bryts sådana snabbt ner till lösa aminosyror.
I den verkliga världen (inte E.s drömvärld) ligger de olika proteinfamiljerna88 utspridda som små planeter på ljusårs avstånd från varandra i den så kallade ”sekvensrymden”, det vill säga på oerhörda mutationsavstånd. Det finns åtminstone fyra andra experimentella studier89 som bekräftar Axes resultat, så det empiriska underlaget är pålitligt.
E.s värld ser som sagt mycket annorlunda ut. I hans universum ligger alla enzymplaneterna i direktkontakt med varandra i en enda stor samling så att det på sin höjd är några enstaka mutationers avstånd mellan dem, eller att de till och med sitter fast i varandra. Därför behövs det bara någon liten punktmutation för att en helt ny funktion kan uppkomma (s 110), och den sortens förändringar är det enda som har behövts från den tid då det (återigen i E.s värld) bara existerade fria aminosyror och några få nukleotider. Men så saknar den också empiriskt underlag, förutom på en ytterst trivial nivå. Jag återkommer till det.
Små enzymmolekyler?
Som vi såg tidigare (s 60) tror E. att nyheter är helt överflödiga under evolutionen. Det räcker med att variera det som redan finns. Många enzym kan ha aktivitet på flera olika kemiska substanser och på det sättet kan miljöförändringar leda till att enzymet blir lite bättre på att ta hand om ett substrat på bekostnad av ett annat längs lång glidande skala av årmiljarder.
E. stöder sin syn på två saker: 1. Små molekyler kan ha katalytiska (enzymlikande) egenskaper, och 2. Nutida enzymer kan skifta substrat genom selektion. (s 109)
För det första är det ytterst långsökt att jämföra de här små molekylerna med moderna enzym. Varför då? Jo, de där små molekylerna kan ha en katalytisk effekt som kanske fördubblar eller möjligen femdubblar reaktionshastigheten för en viss kemisk reaktion. Men enzym i begreppets sanna bemärkelse ökar reaktionshastigheter med faktorer som miljoner eller miljarder! Det är skilda världar även här.
E. föreställer sig en selektion av korta aminosyrakedjor redan innan livet fanns. Men selektion kan inte existera innan självreproducerande organismer redan finns på plats, så det är ett våldsamt tankeskutt mellan de där små molekylerna och ett verkligt enzym. Vetenskapen känner inte till, och har inte ens någon ledtråd till hur de skulle kunna bli till utan cellulära informationssystem och komplexa cellmaskinerier. Eller design förstås.
Det existerar inget molekylärbiologiskt stöd för föreställningen om en värld av livsformer utan de mycket stora enzymer som katalyserar livets grundläggande reaktioner som till exempel replikationen (kopieringen) av DNA
Att en levande primitiv cell skulle kunna existera utan nuvarande integrerade samspel mellan fortplantningssystem, ämnesomsättning, energisystem, materialtransportsystem med mera är rena papperskonstruktioner som också saknar stöd i evidensen. Det är inte förbjudet att spekulera om hur små molekyler successivt kan ha bildat små primitiva system av växelverkande molekyler som kom att integreras med varandra och som med tiden blev så komplexa som de som kan observeras idag. Men då ska det tydligt framgå att det är rena spekulationer och ingenting annat. Någon sådan reservation hittar jag inte i boken.
Visserligen använder sig E. av uttryck som ”vi vet inte”, ”inte alls orimligt”, ”inte alls osannolikt”, ”skulle kunna”, ”skulle vara möjligt”, ”det kan räcka med” i det här kapitlet, men ännu oftare formuleringar som ”behöver bara en mutation”, ”det vi faktiskt vet visar”, ”måste ha varit”, ”undan för undan blivit”, alltså antydningar om att vetenskapen skulle ha full koll på det här.
Nutida enzymer kan skifta substrat genom selektion
Ja, enzymer är mycket riktigt formbara, precis som E. säger (s 110). Men det finns gränser. Vi har redan sett att ett tiotal mutationer som regel räcker för att helt slå ut funktionen hos ett enzym.
Jag och E. är helt överens om att när ett enzym har en viss aktivitet på ett antal substrat i en varierande grad, så kan aktiviteten ökas eller minskas genom mutationer och selektion, ungefär som i exemplet med Dawkins datorsimulering. Det finns mängder av exempel på det, och det är inget som han egentligen behöver nämna. Det E. däremot skulle behöva presentera evidens för är att ett enzym genom mutationer och selektion förvärvar en aktivitet på ett substrat som det helt saknar. Det finns ett massivt experimentellt stöd för att en svag befintlig aktivitet på ett substrat är en förutsättning för att denna ska kunna ökas genom selektion. Återigen handlar det inte om survival of the fittest, utan arrival of the fittest.
Fitnesslandskap
Ett annat sätt att åskådliggöra – och problematisera – saken är med hjälp av begreppet fitness-landskap. Det var ett begrepp som introducerades av den amerikanske genetikern Sewall Wright nån gång på 1930-talet.90 Han tänkte sig ett kuperat landskap fullt med berg och dalar. I det landskapet tänkte han sig att populationer av levande varelser rör sig över tid i takt med att deras gener förändrades genom mutationer. Högt belägna punkter motsvarade hög fitness, det vill säga populationer som var väl anpassade till sin livsmiljö och dalarna sådana som var mindre lyckligt lottade, det vill säga de med sämre reproduktiv framgång (färre ungar). I det här scenariot såg det naturliga urvalet till att organismerna tenderade att alltid röra sig mot högre terräng (till exempel de som råkat få mutationer som gav dem gener för ett lite mer effektivt enzym). Han tänkte sig sedan att landskapet förändrades över tid så att det inte alls var säkert att en organism som nått toppen skulle förbli där för alltid. En miljöförändring skulle när som helst kunna förvandla bergstoppen till en dal och då började selektionen mot en ny topp.
Att tänka i termer av fitnesslandskapet är ett bra sätt att förstå evolutionära mekanismer, men på samma gång avslöjar det ett allvarligt problem med evolutionsteorin. För det första: om organismen hamnar på ett område med slät mark, så har det ingen chans att hitta någon riktning, utan det blir ett slumpartat irrande. Risken är då betydligt större att de vandrar i en riktning som leder bort från det högsta berget i landskapet än mot det. Det motsvarar en organism med ett enzym som helt saknar aktivitet (effekt) på ett visst substrat, för det kan ju inte finnas någon drivkraft att effektivisera en egenskap som inte existerar.
För det andra: problemet med lokala höjdpunkter i landskapet. Om alla varelser söker sig uppåt i terrängen så är ju chansen liten att de råkar befinna sig vid foten av just det högsta berget. Chansen är betydligt större att den närmaste höjden är en liten kulle eller jordhög. Då kommer det naturliga urvalet leda dem upp till toppen på den lilla högen. Väl där kommer fortsatta mutationer bara resultera i att den aktuella egenskapen försämras, och de individerna kommer att elimineras av det naturliga urvalet så att de förblir på den lilla jordhögen. Ju högre den lilla högen är, desto fler lämpliga mutationer erfordras det för att de ska komma ner från höjden och börja utforska en annan del av landskapet. Det här brukar i den evolutionistiska litteraturen kallas ”problemet med lokala fitnessmaxima” eller ”peak-shift-problemet”.
Men det tycks inte bekymra E. Han håller fast vid sin föreställning att små förändringar kan plussas på och skapa människor och makrillar från en primitiv bacill, eller rent av från en pöl med lite aminosyror med ”katalytisk aktivitet”. (s 109)
De här missförstånden av fundamentala och empiriskt välbelagda proteinkemiska förhållanden är själva grunden för E.s gradualistiska syn som återkommer vid upprepade tillfällen i boken. Ett citat från början av kapitlet får illustrera det.
Evolutionsteorins logik bygger på att de tidigaste livsformerna måste ha varit enkelt uppbyggda, och att de undan för undan blivit mer komplexa och utvecklats till de livsformer vi ser idag. Det gäller även proteinerna. (s 108)
Han tillägger att de ”torde” haft
någon form av system för ärftlighet …. och … ämnesomsättning … [med] fungerande enzymer. (s 108)
Så sant. Utan fortplantningsförmåga och ämnesomsättning – inget liv per definition. Det här är verkligen ett lackmustest på hur trovärdigt och empiriskt förankrat den evolutionära berättelsen är. Vi ska därför slutligen ställa frågan: vad är den utifrån observationer och experiment minsta tänkbara rekvisita som krävs för att vi ska ha någonting som ens kan kallas liv?
Vi har idag en ganska god uppfattning om detta, baserad på empirisk vetenskap. Den metod som har använts är följande:
Man har utgått från den enklast byggda naturliga organismen vi känner till, en bakterie med det vetenskapliga namnet Mycoplasma genitalium. Sedan har man tagit bort så mycket DNA man kan utan att bakterien dör eller förlorar förmågan att fortplanta sig. Vetenskapen känner inte till något mer primitivt liv än detta.91
Vi är då nere på 473 gener. 30 procent av dem har ännu okänd funktion, men är ändå nödvändiga för överlevnad. De består sammanlagt av 531 000 nukleotider (DNA-bokstäver). Antalet olika enzym räknas i hundratal, vissa bestående av långt över tusen aminosyraenheter. Läsaren kan enkelt skaffa sig en överblick över all den övriga rekvisita som den enklaste kända livsformen behöver.
Naturligtvis kan man som E. spekulera i att livsformer kan vara enklare än så, och att dessa utvecklats steg för steg under årmiljonerna mot allt mer komplexa organismer. Men då ska man i ärlighetens namn tala tydligt om att nu har vi lämnat den empiriska vetenskapen och befinner oss i rena spekulationer. Det gör inte E. Tvärtom. Han framställer saken som att vi skulle ha goda vetenskapliga skäl för att tro på den där gradvisa processen.
”Detaljen” E. utelämnade
Det sista jag vill påtala är att E. avsiktligt utelämnar en viktig del av ID:s kombinatoriska argument i sin bok. Han urskuldar sig genom formuleringen:
Observera att jag här inte diskuterar sannolikheten för att själva aminosyrorna eller nukleotiderna skall kunna bildas… (s 109)
Det är ett genomtänkt beslut för att slippa reda i ett monumentalt problem för varje livets-ursprungs-scenario, men det är samtidigt en metod att försvaga det kombinatoriska argumentet. Det behandlas av både William Dembski och Stephen Meyer, till exempel Signaturen i cellen på sidan 191-192 (i engelska versionen på s 205-207). Det handlar om en egenskap hos flertalet av de molekyler som ingår i levande varelser, nämligen någonting som kallas kiralitet. Det är värt ett eget kapitel. Problemet med kiralitet hör man nästan aldrig talas om när forskare presenterar fynd av aminosyror i abiogenesexperiment eller i meteoriter, och E. låtsas inte heller om att det existerar. Men det är centralt. Läs om det i Appendix 2.
E. skriver i slutet av kapitlet att den kunskap vi har idag tyder på att slump och evolutionära processer är tillräckliga som förklaring till hur de första enzymmolekylerna uppstod (s 111).
Man kan inte göra anspråk på att vara vetenskaplig om man förnekar eller inte låtsas om kemiska realiteter. Och i nästa andetag beskylla meningsmotståndarna för att bygga sina resonemang på okunskap och halmdockor. Kort sagt – beträffande det kombinatoriska argumentet har ID evidensen helt och hållet på sin sida medan E. hittar på sagor och för läsarna bakom ljuset.
De argument som E. använder som stöd för sin tes saknar alltså stöd i evidensen. De är – med hans egna ordval – ”argument från okunnigheten”.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 17. Det sjätte argumentet: Informationsargumentet
Först lite information om information
Jag har redan nämnt det biologiska informationsbegreppet många gånger i det här dokumentet. Det beror på att det är så centralt inom de biologiska vetenskaperna. Det är inte realistiskt att behandla informationsfrågan på djupet i den här artikeln, och jag är heller ingen utbildad informationsteoretiker, men man behöver inte vara det för att kunna förstå hur relevant informationsfrågan är med avseende på frågan om vårt ursprung, och vilken nagel i ögat den utgör för ateister och agnostiker.
E. menar dels att ID-rörelsens informationsargument är meningslöst eftersom deras variant av information inte går att mäta (s 117). Däremot finns det, menar han, en annan variant som verkligen går att mäta, och det är den enda som är relevant (s 114). Vi behöver därför ta en stund och reda ut det här med informationsbegreppet.
Olika slags information
Förenklat kan man säga att det finns olika nivåer av information. De följande tre bokstavsföljderna får illustrera detta:
ldjfri wnfjfiqpw kntyqqgybybh p hhybggfwäidslgfhåpnaprgönhoivb dktowäqmcöegjji (1)
ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab ab (2)
Tänk dig till sist en följd av bokstäver som den mening du läser i just detta ögonblick! (3)
Den första bokstavsföljden är slumpmässig, den andra lag-/regelstyrd och den tredje skapad av mänskligt medvetande (”intelligens”).
Ospecificerad information
Den amerikanske matematikern och elektronikingenjören Claude Shannon (1916-2001) brukar betraktas som informationsteorins grundare. Han formulerade en matematisk modell för mängdbestämning av informationsinnehållet i en godtycklig följd av symboler. Modellen bygger på sannolikheter - ju mer osannolik en viss symbol är, desto större informationsinnehåll bär den. Men informationsinnehållet är detsamma oavsett om bokstavsföljden är helt meningslös eller djupt meningsfull, och det gör att mängden ”Shannoninformation” inte fångar det som är mest relevant och intressant, det vi i dagligt tal menar med information – nämligen meningsfullhet – den egenskap som särskiljer den tredje bokstavsföljden från de båda andra92.
Graden av komplexitet fångas av begreppet Kolmogorovkomplexitet. Det fångar skillnaden mellan exempel 1 och 2 och säger att 2 har en lägre komplexitet än 1 därför att det uppvisar ett mönster.
Designteoretiker93 skulle säga att symbolföljden (1) ovan är ospecificerad, men väldigt komplex94. Den motsvarar så kallat kosmiskt brus, flimret på en kanallös TV, eller gasmolekylernas rörelser i luften.
Lagstyrd information
Nummer (2) präglas av ett upprepat (eller ”repetitivt”) mönster, likt det man finner i form av vågmärken på en sandstrand, radiosignalerna från pulsarer i rymden, vattenvirveln i ett badkarsutlopp eller så kallade fraktaler inom matematiken. Det är den sortens mönster som våra naturlagar åstadkommer. Även om mönstret är specifikt (i bemärkelsen att det passar in i ett bestämt mönster) så är det inte komplext, eftersom det går att komprimera till en liten banal formel som i det givna exemplet lyder n•(ab), där n=28.
Många naturliga företeelser karakteriseras av kombinationer av slumpmässighet och lagbundenhet, som t ex vädersystem, havsströmmar och den regelbundna strukturen hos en snöflinga.
Meningsfull information
Bokstavssekvensen (3) är den enda av de tre som med rätta kan kallas för en ”mening”. Intelligenta varelser som du och jag har förmågan att producera symbolföljder som bär meningsfull information från en avsändare (i det här fallet mig) till en mottagare (i det här fallet dig som läsare) med ett bestämt syfte (här: att bli lite mer informerad om ämnet för den här texten). Sekvensen är specificerad därför att den är både språkligt och innehållsmässigt relevant, begriplig och meningsfull, det vill säga funktionell. Den är dessutom komplex, i likhet med nummer (1).
Designteoretiker menar att det är just den här informationsnivån som präglar levande varelser. De menar att just denna nivå alltid har ett intelligent upphov. Varje ögonblick skapar intelligenta, medvetna varelser bevisligen stora mängder information som är både specificerad och komplex. Via induktion menar de att detta utgör ett starkt argument - eller ”bevis” om man så önskar - för att det finns en intelligent Skapare som orsak till jordens livsformer; en aspekt av det teologer kallar ”den allmänna Gudsuppenbarelsen”. Just den här tredje och sista egenskapen hos information är det som är i fokus här och nu. Det är också just det som E. försöker tona ner eller rentav eliminera betydelsen av.
I sin bok Signaturen i Cellen resonerar Stephen Meyer ungefär så här:
Funktionellt specificerad information i cellen
Det är känt sedan 1950-talet att DNA-molekylen är livets universella informationsbärare. Molekylen förekommer i alla kända livsformer.95 Det är den inbördes ordningen (sekvensen) av nukleotiderna längs med DNA-molekylen som bär informationen, på samma sätt som bokstäverna i den här texten gör det. Det finns inga kemiska eller fysikaliska lagar som bestämmer ordningen på dem.
• Cellen innehåller bevisligen funktionellt specificerad digital information.
• Cellen innehåller dessutom bevisligen funktionellt integrerade system för att behandla denna information.
Digital information i cellen?
På vilket sätt anser då Meyer att cellen innehåller digital information?
Informationen är digital därför att den likt talade språk, morsekod och datorspråk bygger på meningsfulla arrangemang av diskreta, avgränsade symboler. De kan exempelvis vara i form av bokstäver, ljud- eller ljussignaler eller ettor och nollor (två olika elektriska spänningar). I morsekod eller datorkod handlar det om binära koder (med två symboler), medan det i DNA är det en kvarternär kod bestående av fyra symboler (de fyra kvävebaserna som förkortas A, C, T och G), men principen är en och densamma: symboler arrangerade i sekvenser förmedlar meningsfulla budskap.
Specificerad information i cellen
Den andel (kanske 1,5 procent)96 av den mänskliga arvsmassan som utgör ”recept” på de proteiner som behövs i människokroppen, det vill säga de proteinkodande generna, är mycket specifika, i synnerhet den tredjedel av dem som kodar för enzymer som hanterar kroppens ämnesomsättning. De kännetecknas av just specificitet, vilket innebär att deras funktion i kroppen är att underlätta (katalysera) en speciell typ av kemiska reaktioner.
Meyer använder sig framför allt av termen specificerad komplexitet, men det är i princip synonymt med funktionell komplexitet.
Funktionellt integrerade system i cellen
Vad menar Meyer då med begreppet funktionellt integrerade system? Han syftar till exempel på det ytterst komplexa molekylära maskineri som möjliggör för den digitala informationen i nukleinsyran DNA att överföras från en kod till en annan. Den digitala informationen i DNA bygger som sagt på de fyra kvävebaserna A, C, T och G. Men speciella enzym överför vid den så kallade transkriptionen utvalda delar av informationen i DNA till en annan nukleinsyra, RNA, där kodspråket i stället bygger på baserna A, C, U och G. Längre fram i processen sker ytterligare en informationsöverföring i de fall då informationen i RNA överförs till en viss sekvens av aminosyror i ett protein förmedlad av en uppsättning andra enzym i enlighet med det som populärt brukar kallas ”den genetiska koden”. Här har vi alltså flera med varandra integrerade informationssystem som är oupplösligt bundna till och beroende av varandra på sätt som Meyer menar är omöjligt att uppnå genom en planlös mekanism som mutationer och selektion.
Hur hamnade informationen i DNA?
Meyers grundläggande fråga är: Hur hamnade informationen på DNA-molekylen? Molekylen som sådan är inte levande och den är inte heller synonym med information – den är ”bara” bärare av informationen som kodar för alla beståndsdelar, monteringen och funktionen hos den organism som den finns i.
Eftersom svaret inte går att hitta i nuet konstaterar han att det ligger i historien. Och svar på historiska frågor når man genom abduktion till den bästa förklaringen. Därför ställer sig Meyer frågan vad det finns för tänkbara alternativa förklaringar till fenomenet med cellens funktionella information. Han konstaterar att det hypotetiskt kan ske genom antingen en naturalistisk selektionsprocess eller genom design.
Som vi minns konstaterade E. att när man argumenterar abduktivt som i det här fallet så är det viktigt att man baserar sitt val av förklaring på evidens. Meyer ställer frågan om det finns någon evidens till stöd för att naturalistiska processer som till exempel mutationer och naturligt urval har visat sig kunna producera komplex funktionell information. Hans svar på den frågan är nej, det finns ingen evidens för det någonstans i den vetenskapliga litteraturen.97
Nästa fråga Meyer ställer är: Finns det vetenskaplig evidens för att intelligenta orsaker (eller aktörer, eftersom det handlar om levande personliga varelser) kan skapa komplex specificerad information i form av meningsfulla sekvenser av symboler. Svaret är ett oreserverat Ja. Det är precis vad intelligenta aktörer (E. använder sig i stället av begreppet ”agenter”) ständigt ägnar sig åt varje gång de/vi tänker, talar eller skriver. Det är inte bara en känd orsak till uppkomsten av detta slag av information, utan dessutom den enda orsaken som vetenskapen känner till.
Meyers slutsats blir att den idag bästa evidensbelagda förklaringen till livets komplexa information som vi finner i DNA-molekylen är en intelligent aktör som i det avlägset förgångna försåg DNA-molekylerna med denna egenskap.
E.s motargument
E. poängterar att abduktion till den bästa förklaringen måste underbyggas av ”solid evidens”. Det gör inte ID menar han. Hans invändningar mot informationsargumentet är följande tre:
1. Eftersom meningsfull information inte går att mäta är det ointressant att diskutera den.
E. menar att eftersom mängden specificerad information inte går att mäta med några metoder så är det ”knappast meningsfullt”, och de är inte
möjliga att använda i vetenskapligt arbete. De används inte heller inom den etablerade informationsteorin, utan är begrepp som enbart åberopas av kreationister och designförespråkare. (s 117)
2. Det är bevisat att informationen i levande organismers arvsmassor kan öka genom genduplikationer.
Enligt den etablerade informationsteorin kan alltså genetisk information öka genom evolutionära processer. (s 114)
3. ID är inte falsifierbar.
Vi tar dem som vanligt i ordningsföljd:
1. Låt oss göra ett tankeexperiment. Den här boken om Intelligent design innehåller 215 sidor. Den har ungefär 2500 tecken per sida. Det blir totalt cirka en halv miljon symboler.
Vi tänker oss nu en annan bok, också den bestående av 215 sidor och en halv miljon symboler. I den här boken finns exakt samma bokstäver, siffror och skiljetecken, men de befinner sig nu huller om buller i en enda röra.
Tror vi att E. skulle tycka att det är någon skillnad mellan de båda böckerna? Förmodligen. Vad är i så fall skillnaden? Går det att mäta denna genom att räkna antalet symboler, alltså Shannoninformation? Nej, för den är exakt lika stor i båda fallen.
Bör vi därför dra slutsatsen att E.s bok är rappakalja? Det gör inte ens jag, i varje fall inte av den här anledningen. E. borde därför vara ytterst motiverad att uppmuntra ID-rörelsens försök att hitta en metod som förhindrar den sortens förhastade slutsatser.
E.s bok motsvaras av en levande organism bestående av tusentals olika proteiner med olika stabila strukturer som ger dem deras specifika och djupt meningsfulla former och funktioner. Den andra boken skulle motsvara samma volym strukturlösa aminosyrakedjor i form av en behållare med en slät, klimpfri soppa utan en enda stabil proteinmolekyl. Hur kan vi veta det? Jo, därför att det, som vi redan nämnt (kap 16), är experimentellt belagt att sannolikheten att en slumpmässig sekvens av aminosyror (motsvarande ett kort protein) bildar en stabil grundstruktur är så sällsynt som en på 1077.
Meningsfullhet som sådan är alltså en realitet som inte kan sopas under mattan. E. vidhåller att det är en falsk analogi när den kopplas till levande varelser, men den idén kan vi också avfärda. Den är alltför uppenbart kopplad till den biologiska världen genom experiment och observationer.
Idén att bara det som går att mäta existerar och spelar någon roll får märkliga konsekvenser. Det är inte bara meningsfullhet utan även företeelser som skönhet, kärlek och empati som är svåra eller omöjliga att mäta. I det evolutionära perspektivet kan sådant därför aldrig förklaras, bara bortförklaras.
Att förklara biologisk information som ointressant ger E. ett alibi att slippa hantera den besvärliga frågan om informationens ursprung. Det är inte så konstigt, för informationsfrågan torde vara den primära utmaningen för evolutionsteorin.
Sedan har E. fel när han hävdar att funktionell information inte går att mäta. Det finns åtminstone en sekulär studie om just detta.98
Det går också att göra en uppskattning av den undre gränsen för mängden information i en varelses arvsmassa. Det kan man göra genom att mäta hur många kvävebaser (”bokstäver”) i DNA som behövs för att koda för alla de olika proteiner som bygger upp organismen, får den att vara livsduglig och kunna föröka sig, och sedan tillämpa Shannons beräkningsmodell.
Bristerna i Shannonmodellen, som E. förordar (s 114), är att varje enskild gen i medeltal kodar för fem (men ibland upp till tiotusentals) olika proteiner med varierande funktion, och dessutom att samma DNA-text, till skillnad från den här artikeltexten, i cellen läses och översätts till proteiner i flera överlappande dimensioner, däribland baklänges. Då blir det en extrem underskattning att bara räkna nukleotiderna i originalgenen i DNA.
Detta gör det rimligt att förvänta att cellernas information är kraftigt komprimerad och avsevärt större än antalet ”bokstäver” anger (6,4 Gbit99 i människans fall). I dagsläget vet ingen med vilken faktor som cellernas informationsinnehåll bör multipliceras, men att använda detta som ett argument för att helt bortse från frågan om informationens uppkomst är givetvis bara ett svepskäl.
2. Hur förklarar evolutionsteoretiker att det genetiska programmet i en urtida primitiv för-bakterie kunnat utvecklas till de formidabla mjukvaruprogram som präglar nu levande organismer? Svaret bygger på att enskilda gener ska ha råkat ut för mutationer som inneburit att dottercellen av misstag fått dubbla eller flerdubbla upplagor av samma information i förhållande till modercellen (s 112). Sedan förutsätter teorin (under förutsättning att organismen inte tar skada av förändringen och elimineras av det naturliga urvalet) att det sker mutationer i de extra kopiorna till dess att någon av dem plötsligt råkar100 ha förvandlats till en helt ny funktionell gen som leder till en helt ny egenskap. Genom denna mekanism menar man att praktiskt taget alla de gener och egenskaper som skiljer oss och allt annat levande från blågröna bakterier har uppstått under ”årmiljonernas” lopp.101
Kan då genetisk information ”ökas genom evolutionära processer” (s 114). Som sagt, ja, men bara i en helt banal bemärkelse. E. skulle kunnat illustrera det genom att kopiera några godtyckligt valda meningar i sin bok och klistra in dem på ytterligare några slumpmässigt valda ställen i syfte att få mer sagt. Alternativt kunde han ha tagit sig en tupplur med ansiktet tryckt mot tangentbordet. Det skulle ha tillfört boken avsevärda mängder Shannoninformation. Men knappast några bättre argument.
3. Allra sist: E. menar att ID inte är falsifierbar. Men ID gör prediktionen att meningsfull information alltid har ett intelligent upphov. Om motsatsen kan bevisas experimentellt skulle det falsifiera designteorin. Det förutsätter att det inte handlar om kopiering av redan befintlig funktionell information (som genduplikationer eller rent kvantitativa resonemang à la E.s) eller simuleringar där ett designat datorprogram och utvald indata – som båda utgör resultat av en intelligent design – innehåller tillräcklig informationsmängd för att förklara informationen i utdata.
Det här borde också kunna användas för att falsifiera evolutionsteorin, åtminstone temporärt. Om det kan visas att meningsfull information undantagslöst bara kan genereras av bevisligen intelligenta processer, så skulle det vara tillräckligt för falsifiering här och nu. Däremot inte med nödvändighet för all framtid – kanske finns det ännu oupptäckta mekanismer som skulle kunna ge en förklaring.
Men kort sammanfattat: Livets information existerar bevisligen. Idag kan den bara förklaras av en intelligent design.
Information och tanke
Hur kan vi förvissa oss om att den sanna bilden av information inte är den som E. försöker ge läsaren? Genom att inse sambandet mellan information och tanke. Stephen Meyer skriver i sin bok Darwin’s Doubt – vi återkommer till den:
Vi kan känna igen tankeprodukter, effekter av intelligent design, i dess distinkta informationsrika manifestationer. (s 398, min övers)
De rader du just nu läser är en informationsrik manifestation. Det är en effekt av en intelligent design (nåja, men det var i alla fall jag som skrev). Det är mina tankar som jag just nu förmedlar till dig. På samma sätt manifesterar informationen i DNA strukturen hos alla livets molekyler och deras inbördes arrangemang, så att vi kan leva.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 18. Det sjunde argumentet: Förklaringsfiltret
William Dembskis förklaringsfilter är ett logiskt resonemang för att detektera design av ett objekt genom att successivt utesluta andra orsaker som slump och andra naturliga förklaringar (naturlagar).
Demski lägger fram två bevis. Först ett induktionsbevis där man utgår från föremål där vi har facit, som en dator. Det finns inga naturlagar som gör att det med nödvändighet spontant uppstår datorer – den är därmed kontingent.
Den är komplex. Men inte bara komplex i den bemärkelse som ett skrotupplag kan vara det, vilket dessutom gör den specifik. Och därmed kan vi med rätta dra slutsatsen att datorn är ett resultat av en intelligent design. Och vi vet att så är fallet. Vi kan testa olika föremål, designade eller inte, och filtret ger oss nästan alltid rätt svar. Det nämnda skrotupplaget är till exempel komplext, men ospecificerat och därför drar vi slutsatsen att högen inte är avsiktligt designad till att se ut precis som den gör (se figuren ovan).
E. tycker att det är problematiskt att förklaringsfiltrets logik skiljer sig från vetenskapligt arbete av följande skäl:
1.Det handlar inte om ett val mellan två konkurrerande hypoteser, utan mellan en mekanistisk och icke-mekanistisk förklaring, (s 119) vilket inte går att göra med vetenskapliga metoder. (se kap 6).
Min kommentar:
Fallet med SETI visar att E. har fel när han påstår att vetenskaplighetskriteriet inte är uppfyllt (kap 12).
2.Det är mekanistiska förklaringar som prövas i filtret, inte designförklaringen (s 119).
Min kommentar:
Ja, eftersom världen vi befinner oss i när vi bedriver vetenskap är just materiell och det är materiella objekt som analyseras.
3.Om vi saknar kunskap att välja behövs det en skräpkorg för det som varken är nödvändigheter eller slump (s 119).
Min kommentar:
Det som varken är nödvändighet eller slump är antingen design eller en kombination av nödvändighet och slump. E. har en poäng här. Men det är ingen skräpkorg som behövs. Det skulle på sin höjd behövas en utgång för företeelser som går att förklara med hjälp av en kombination av slump och naturlagar (se s 61). Därefter återstår designalternativet. Där hamnar med nödvändighet saker som smartphones, bilar och DNA-molekyler som varken kan förklaras med slump, naturlagar eller kombinationer dem emellan. Så när E. ersätter design med en skräpkorg får han minst sagt problem – det är inte bara DNA som hamnar där. Det gör bilen och smartphonen också.
Men det finns också praktiska problem, menar E., om man vill undersöka om en biologisk struktur är designad eller inte:
1.Hur bedömer man gränsen mellan hög och låg sannolikhet för ett objekt. På vad ska vi beräkna sannolikheten? (s 120)
Min kommentar:
På sådant som vi vet av erfarenhet att slump och naturlagar kan åstadkomma.
2.Det går inte att beräkna sannolikheten på en evolutionär process (s 120)
Min kommentar:
Filtret är tänkt att användas på det fysiska resultatet av processen eller processerna, inte dessa som sådana.
3.Filtret behandlar bara slump eller nödvändighet, men inte kombinationen av de båda. (s 120)
Min kommentar:
Om sannolikheten för kombinationen av de båda är nästintill 100 procent blir det nödvändigt och filtret kommer att fungera ändå.
Är då flagellen osannolik, undrar E.:
Eftersom evolution är den accepterade förklaringen borde man ställa upp den mest sannolika evolutionära hypotes man kan formulera utifrån dagens kunskap och analysera den. (s 122)
Han prövar hypotesen att flagellen har uppstått från lösa aminosyror av en ren slump, inte hypotesen att flagellen har uppstått genom evolutionära processer. (s 122)
Min kommentar:
Som vi såg i kapitlet om det kombinatoriska argumentet så tänker sig E. att det finns en obruten linje från urorganismen till oss och allt annat levande – ja, till och med från fria aminosyror och nukleotider (s 109) – via slump (mutationer) och nödvändighet (naturligt urval). Eftersom han i princip betraktar denna evolutionära process som ett faktum, så kommer utfallet i förklaringsfiltret att bli alternativet nödvändighet. På det viset slipper han bekymret att filtret leder fram till slutsatsen design. E.s resonemang bygger inte på empiriska resultat och evidens, utan utgör en tro eller filosofi som närmast kan jämföras med naturmagi. Naturligtvis är han helt fri att ge uttryck för vilken sorts religion han vill. Det olyckliga är att han förkläder den till vetenskap.
***
Sannolikheten för den evolutionära processen är alltså inte alls hundra procent, utan så nära noll man kan komma. Kom ihåg att åtminstone fem oberoende empiriska studier har bekräftat att proteiner, inklusive bakterieflagellens, inte går att nå genom en selektionsvandring genom sekvensrymden (se kap 16 ovan).
Personer som inte kan tänka sig design som alternativ kan förstås alltid försöka komma förbi detta på olika sätt. Ett är att hävda att om det finns tillräckligt med tid så blir det osannolika sannolikt. Ett annat är – som E. är inne på (s 48, 149) – att det finns ofantligt många fler världar än vår egen (enligt strängteorin kanske 10500). Det som verkar osannolikt i en värld blir sannolikt, eller nästintill nödvändigt i en annan, som råkade vara vår.
Granskningen av det kombinatoriska argumentet (kap 16) visade att E.s kritik bygger på en kombination av en övertro på selektion och evolverbarhet och Dawkins missförstånd med vesslan – det som utmålades som en naturlig selektionsprocess men som i praktiken var en intelligent designprocess. Design blir därför den enda rimliga slutsatsen.
Enligt E. förhåller det sig så, att om man inte accepterar den evolutionsbiologiska förklaringen
skulle designern personligen behöva tillverka varje enskild flagell i varje cell hela tiden. och detta sedan miljarder år. (s 122)
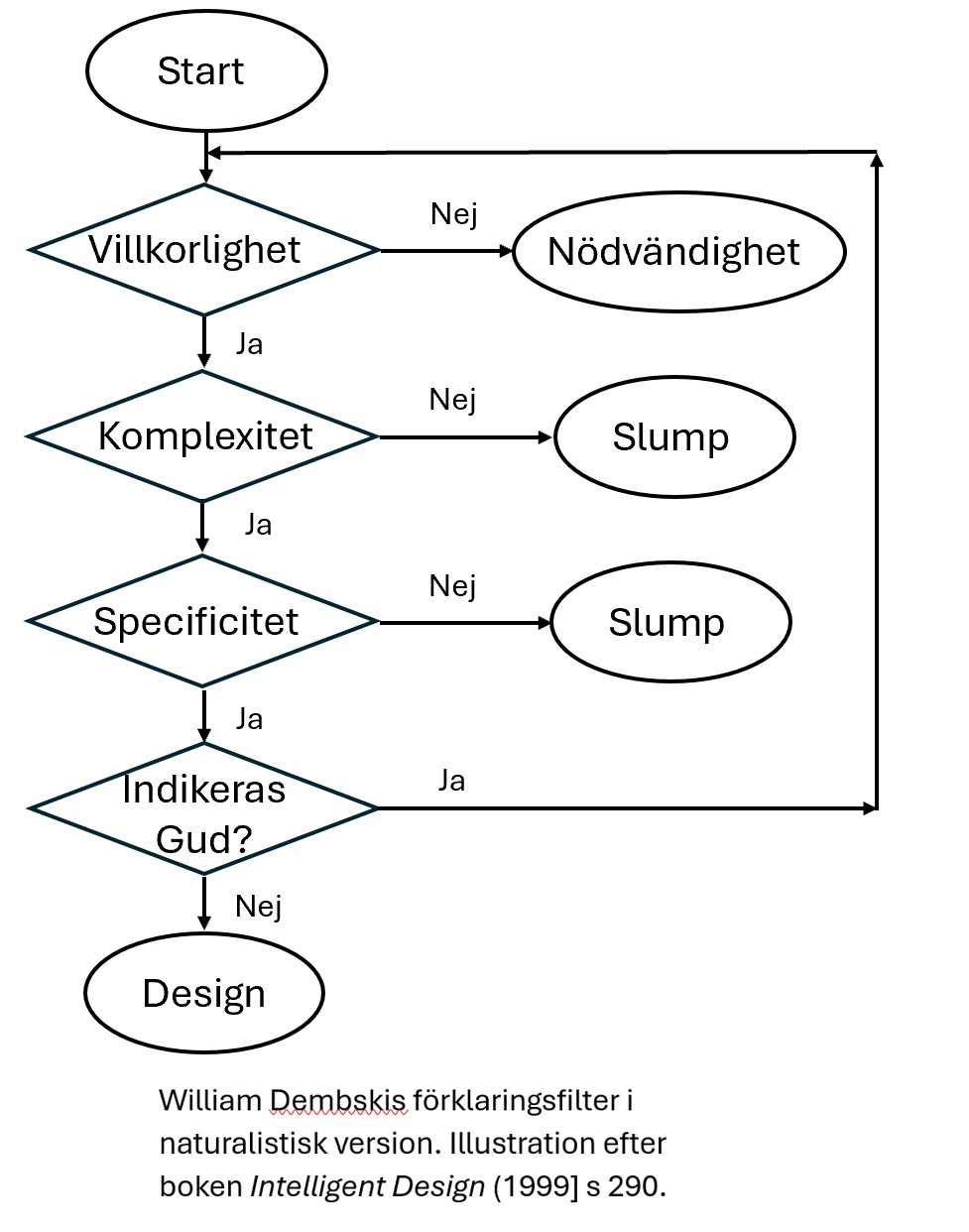
Men det här är i bästa fall ogenomtänkt av E.. Proteinerna i en flagell och de instruktioner som erfordras för deras montering är varken mer eller mindre specifika än det DNA som kodar för dem. När koden för deras struktur väl existerar kan cellens mekanismer både föra den vidare till nästa generation (replikation) och förverkliga dem genom att tillverka de aktuella proteinerna (translation).
Till sist: Jag är benägen att hålla med E. om att det inte är så lätt att med nuvarande kunskapsläge tillämpa förklaringsfiltret med matematisk stringens på strukturer i den levande världen därför att dessa innehåller storheter som är svåra att kvantifiera. Men det kanske inte heller behövs. Empiriskt baserade uppskattningar kan räcka långt, och som en generell modell är Dembskis förklaringsfilter både elegant och intuitivt begriplig. Det handlar mer om att vilja förstå argumentet. Det gör inte E. Det är alltid oklokt att kasta ut barnet med badvattnet även om man inte skulle tycka om det.
William Dembski förutsåg E.s invändning och skapade även en naturalistisk variant av sitt förklaringsfilter som effektivt utesluter Gud som Designer (se illustrationen ovan).
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 19. Det åttonde argumentet: Skönhetsargumentet
Evolutionära förklaringar av skönhet
Naturens skönhet är alltför påtaglig för att kunna bortförklaras av evolutionister, alltså behöver den förklaras – utan en Skapare. Hur går det till?
E. presenterar i det här kapitlet två olika förklaringar till naturens mångfald av skönhet med avsikt att ta udden av det som designargument. Den ena är neutral evolution och den andra sexuell selektion.
Neutral evolution är i princip mutationer utan selektion. Mot bakgrund av vad som är sagt om mutationernas nyskapande förmåga så borde det inte finnas så mycket att tillägga. Om kombinationen mutationer + naturligt urval är en minst sagt svag förklaringsmodell (i praktiken dubbelt destruktiv), så torde ett scenario där ”nära-neutrala” mutationer skapar nya anlag för nya egenskaper utan att selektionen skapar en riktning för förändringarna närmast vara en ren utopi. E. framställer det mer eller mindre som ett bevisat faktum. Det är:
en etablerad del av evolutionsteorin i ett halvt århundrade. (s 125)
Men evidensen lyser med sin frånvaro. Den utgörs av en obevisad hypotes i form av en evolutionär berättelse där saker ”råkar” bli till:
[D]et är just det vi skulle vänta oss av en neutral evolution; alla detaljer i ett lövs uppbyggnad behöver inte ha någon tydlig funktion, de kan bara råka finnas där. (s 125)
Så den sexuella selektionen. Det har under historien lagts fram olika hypoteser för att förklara saker som påfågelns fjäderdräkt eller lövsalsfåglarnas parningsritualer. Den vanligaste bland evolutionsbiologer är att (oftast) honor väljer att para sig med de snyggaste makarna så att varje ny generation blir aningen snyggare till dess att skönheten är närmast fullkomlig. E. presenterar evidens från en kollega vid Göteborgs universitet som skrivit en artikel om att det fungerar så i praktiken. (s 126)
Det där låter som en logiskt förklaringsmodell, men det finns evidens som talar i en annan riktning. Jag har lagt en längre diskussion om fenomenet skönhet i Appendix 3 för den som är intresserad av att fördjupa sig.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 20. Det nionde argumentet: Kambriska explosionen
Kapitlet om den kambriska explosionen är bokens längsta kapitel och upptar hela tolv sidor. Vad menas med det argumentet?
Det är det välkända och etablerade faktum att det i ett visst berglager, som sekulära forskare menar lades ner för mellan 530-525 miljoner år sedan uppträder ett stort antal väldigt olika organismgrupper i fossilen som inte går att påträffa längre ner i de fossilförande lagerföljderna (s 127). Namnet kommer av att det tycks som om livet bildligt talat exploderade just då. Det handlar därmed om en period på en promille (tusendel) av jordens (förmodade) evolutionära historia.
Olikheterna bland organismerna brukar beskrivas som att de har kombinationer av grundläggande byggnadsplaner (arkitektur) som särskiljer dem från varandra. Man säger att de tillhör olika fyla (singular fylum), på svenska ”stammar”.
Stephen C. Meyer har skrivit en bok om den kambriska explosionen, Darwins Doubt, där han argumenterar för att den stora biologiska mångfald som uppträder så plötsligt i lagren måste höra samman med en motsvarande ökning i mängden funktionell information. Meyer nämner speciellt de leddjur som kallas trilobiter som var försedda med ytterst avancerade komplexögon med förmåga till 360-gradigt bildseende.
Meyer noterar särskilt det udda förhållandet att det är just på en så hög hierarkisk nivå som fyla som mångfalden uppträder. Av en gradvis evolution skulle man förvänta sig att de fåtaliga organismtyper som föregick de kambriska skulle förändras långsamt och över tid ge upphov till nya underarter, så småningom nya släkten och därefter under långa tidsrymder nya familjer, ordningar, klasser och till sist nya fyla ungefär som ett träds stam, grenar, kvistar och skott. Det är också den bild som E. beskriver (s 131).
Men det verkliga förhållandet är som sagt det omvända – fyla är den högsta hierarkiska nivån under de olika rikena, och de dyker upp längst ner i de sedimentära berglagren. Evolutionen tycks alltså ha börjat med fyla och slutat med underarter. Det rimmar väldigt dåligt med evolutionära förväntningar menar Meyer. Det evolutionära trädet tycks alltså på sätt och vis vara upp-och-ned. Det är splitter nya fyla som dyker upp i de kambriska lagren och först högre upp i lagren tycks de ha differentierats i olika familjer, släkten och arter.
De förslag från evolutionistiskt håll för att hantera det här problemet har varit hypotesen att det skedde förändringar i vissa regulatoriska gener som ledde till alla de här drastiska omstruktureringarna av djurens byggnadsplaner. Problemet med en sådan hypotes är att det inte finns någon experimentell evidens som stöder idén. Den evidens som finns pekar entydigt mot att förändringar i den sortens gener (till exempel hox-gener) undantagslöst leder till allvarliga missbildningar eller död. Det är sådana mutationer som genererar bananflugor med ben på antennernas plats eller fyra vingar i stället för två (där två inte går att använda och flugan knappt kan flyga).102
Kom ihåg det nästintill oöverstigliga problemet för mutationer att generera fler än två ändamålsenliga mutationer i ett protein utan att förstöra funktionen längs vägen (se diskussionen om Michael Behes studie på malariaparasiten (kap 15 ovan). I ljuset av det förefaller det såväl teoretiskt som praktiskt uteslutet att ens något enda av alla de hundratusentals splitter nya proteiner (och gener) som behövs för den kambriska faunan skulle kunna uppstå inom ramen för universums sekulära ålder. Och då har vi inte ens berört frågan om hur den epigenetiska programmeringen (se kap 8 ovan) som är helt avgörande för embryons utveckling.
Konsten att bli kvitt en explosion
E. vill vederlägga den kambriska explosionen som ID-argument. Låt oss se hur han resonerar:
Först och främst vill han bortdefiniera problemet genom att hävda att det bara existerar i huvudet på designanhängare:
Man ska dock vara medveten om att fyla, liksom alla andra systematiska kategorier, inte finns i naturen utan bara i våra huvuden. (s 128)Man bör också vara medveten om att fyla inte representerar helt unika kroppsplaner, utan att de har inbördes likheter som gör att de kan inordnas i överordnade kategorier på ett sätt som visar en trädstruktur med successiva förgreningar. (s 128)
Det här är direkt missvisande. Fyla definieras just av grundläggande kroppsplaner. Det förhållande att vissa drag kan delas mellan olika fyla förändrar inte den saken. Den läsare som tror att jag fabulerar kan rådfråga sin AI-assistent. Jag ställde själv frågan till ChatGPT:
Vad är din kommentar till följande påstående: ”Fyla finns inte i naturen utan i bara våra huvuden”?
ChatGPT svarade:
Påståendet är överdrivet och i praktiken fel. Även om fyla är mänskliga klassifikationer återspeglar de verkliga, djupa evolutionära skillnader i kroppsplan som faktiskt finns i naturen,
Observera att det fetstilta är ChatGPT:s eget val. Referensen till evolutionen beror naturligtvis på att roboten är djupt indoktrinerad i ett evolutionärt tänk. E. bör följaktligen ta en djupdiskussion med sin egen assistent och se ifall han kan lyckas övertyga den. Jag betvivlar det starkt.
När E. nu ägnat en god stund åt att förklara att begreppet fylum bara är en tankekonstruktion utan någon direkt koppling till verkligheten går han raskt vidare med att försöka visa att dessa nog-icke-existerande kategorier:
1. har en mycket stor inbördes variation. (s 128-131)
2. bara i liten, eller kanske bara skenbar omfattning återfinns i kambrium. (s 132-136).
E.s poäng med den första punkten är att det inte spelar någon större roll om man hittar ett ryggsträngsdjur i prekambrium, för de är ju så små och enkla i relation till grodor, människor och elefanter.
Poängen med punkt två är att avdramatisera den kambriska explosionen som ett problem för evolutionsteorin eftersom det enligt honom hände minst lika mycket både före och efter denna ”period”. Att det skenbart tycks ha hänt så mycket i just kambrium, menar han, är att det blev mer syre i atmosfären så att djuren kunde bli större och att det blev mer kalcium i haven så att djuren kunde börja bilda skelett och skal. Och nu började en del djur få bildseende tillägger han. (s 137)
I figur 11 (s 135) illustrerar E. de ”tidpunkter” när de olika fyla uppträder. Bortsett från ett mindre antal fyla som han märkt ut lite ”halvvägs mellan kambrium och nutid, så ligger de allra flesta samlade kring någon av dessa båda ändpunkter. Det finns fossila spår i kambrium efter merparten av de mellanliggande grupperna.103
Det intressanta är att de djur som utan minsta tvivel återfinns i den kambriska faunan är makroskopiska sådana, det vill säga sådana som är stora nog att se med blotta ögat. De som i E.s diagram ligger långt till höger, och som alltså enligt honom utvecklats först på senare tid, är med få undantag mikroskopiskt små inälvsparasiter hos ryggradsdjur. Det vill säga organismer som vi knappast skulle förvänta oss att hitta som fossil.
E. vill med diagrammet visa på hur djurlivets evolution ägt rum kontinuerligt under 600 miljoner år. Samma datauppsättning kan tolkas som att alla fyla existerade redan i kambrium, men man har bara påträffat de av dem som haft en sportslig chans att bevaras där som fossil. Perspektiv gör skillnad.
Artefakthypotesen
E. driver den så kallade artefakthypotesen, även om han inte nämner den vid namn. Det är en hypotes som tagits fram för att bortförklara det faktum att det generellt sett helt saknas föregångare i lägre belägna lager till alla de livsformer som dyker upp i kambrium. Hypotesen tar sig lite olika former, men den vanligaste är helt enkelt att alla de berglager som dokumenterat denna långa evolution har försvunnit och inte längre går att påträffa.
En annan vanlig variant är att man letat alltför dåligt efter fossilen så att de luckor man ser bara är skenbara, man kommer att hitta övergångsformer bara man fortsätter leta,
Ytterligare en variant är att förmågan att bilda skal och skelett och stora djur uppstod först i kambrium genom att havets kemi kom att innehålla mer kalcium och syre. Det var först nu som de kunde bevaras som fossil, hävdar man.
Den första varianten är inte så mycket att säga om. Den är och förblir just en bortförklaring.
Den andra varianten motsägs av statistiska undersökningar.104 Principen är att trots att man ständigt hittar nya fossil så faller dessa praktiskt taget undantagslöst i just de systematiska kategorier (släkten familjer, fyla etc) som redan finns. Varje nytt fynd som görs bekräftar därmed att de befintliga kategorierna är verkliga kategorier.
Den tredje varianten motsägs bland annat av att det yttre skelettet hos leddjur (som trilobiter) eller armfotingar utgör en integrerad del av djurens anatomi. Det kan handla om sådant som anslutningar till muskelsystem för rörelse, system för skalömsning som möjliggör djurens tillväxt och mycket annat.
E. är mycket kritisk till Meyer. Han skriver:
Han [Meyer] läser in hierarkiska strukturer i naturen och menar att de påminner om människans sätt att organisera system i hierarkier. Men analogier bevisar ingenting… (s 138)
Men E. vittnar mot sig själv. Han har tydligen glömt att han tidigare i kapitlet förklarat hur det kom sig att ryggsträngen hos ryggsträngsdjuren behölls under evolutionen med följande motivering:
Ryggsträngen är så basal och så integrerad i embryonalutvecklingen att den svårligen går att avlägsna. Detsamma gäller de genetiska nätverk som styr den tidiga utvecklingen. (s 131)
Men vad är det E. här beskriver om inte just den hierarkiska struktur som han sju sidor senare beskyller Meyer för att helt obefogat ”läsa in” i naturen?! Nån konsekvens får det allt lov att vara. Han fortsätter:
Dessutom beskriver man idag biologiska system som nätverk snarare än hierarkiska system. (s 138)
Som så ofta presenterar E. här ännu en halvsanning (eller snarare halvlögn beroende på vilket humör man är). En sanningsenlig beskrivning av dagens kunskapsläge är att biologiska system är hierarkiskt organiserade men nätverksmässigt integrerade. Det handlar alltså inte alls om att man övergett en syn till förmån för en annan. Vem av oss båda talar sanning? Återigen – fråga din assistent eller googla.
Man behöver inte vara systembiolog för att inse det felaktiga i E.s resonemang. Vi vet alla att små biomolekyler (t ex aminosyror och nukleotider) bygger upp makrobiomolekyler (som DNA, RNA och proteiner) som bygger upp organeller (ribosomer, mitokondrier…) som bygger upp celler (hudceller, leverceller, muskelceller…) som bygger upp vävnader (fettvävnad, muskelvävnad…) som bygger upp organ (hjärta, njurar…) som bygger upp organsystem (andningssystem, cirkulationssystem, matspjälkningssystem…) som bygger upp organismer (bananplantor och människor) som bygger upp ekosystem som bygger upp jordens biosfär. Vad är detta om inte just en hierarkisk organisation?105 Strukturerna på varje nivå äger egenskaper och funktioner som underliggande nivåer saknar. Funktionerna uppstår genom det sätt på vilket delarna på varje nivå är arrangerade och hur de samverkar med varandra.
Stephen Meyers huvudsakliga argument i boken är att alla de nya byggnadsplaner som uppträder i de kambriska lagren kräver att stora mängder med ny biologisk information som är strukturerad på unika sätt i de olika kategorierna av djur (fyla) uppträder på mycket kort tid.106 E. invänder:
Emellertid är det omöjligt att veta hur mycket information som skulle behövas, eller hur snabbt den skulle kunna uppstå. (s 138)
E. avfärdar alltså behovet av ny genetisk information som grundlöst med argumentet att det inte går att beräkna mängden av information och den hastighet med vilken den måste ha uppkommit och med att Meyer inte redovisat någon beräkningsmodell. Vi har hört det förut (se kap 17 ovan). Hans budskap är entydigt: Det som inte går att mäta och beräkna existerar inte.
Låt oss då dra slutsatsen att evolutionsteorin inte heller existerar, för vem kan väga eller mäta den?
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 21. Det tionde argumentet: Väntetidsargumentet
Låt oss tänka oss följande scenario:
En viss man råkar födas med en fördelaktig mutation, låt oss säga en utseendemässig egenskap som en hårtofs på näsan, som av kvinnor uppfattas som mycket attraktiv. Hur lång tid kommer det att ta innan jordens hela befolkning har denna gen så att alla män har hårtofsen i fråga på sina näsor (populationsgenetiker säger då att genen blivit ”fixerad” i populationen)?
Vi inser genast att det kommer att ta mycket lång tid, i synnerhet om befolkningen är stor och om anlaget är recessivt, det vill säga inte framträder förrän en man ärver det från båda sina föräldrar. Detta är anledningen till att evolutionsbiologer spekulerar om att populationen inte var större än ca 10 000 när människan utvecklades.
Det här är väntetidsargumentet. Det är nära förknippat med det så kallade Haldanes dilemma, som innebär att selektionen alltid har ett högt pris i form av minskad genetisk mångfald i och med att genvarianter elimineras längs med vägen.
Datorberäkningar understryker att den insikt som det nämnda exemplet ger också är förankrat i den genetiska verkligheten. De visar att den tid som krävs för en enda fördelaktig punktmutation att bli fixerad i en mänsklig population på 10 000 individer107 är i genomsnitt 12 miljoner år – det vill säga dubbelt så lång tid som den hypotetiska omvandlingen av ”apa” till människa. Och redan en skillnad på 1-2 procent mellan människa och schimpans (som i praktiken är betydligt större) innebär 30 – 60 miljoner DNA-bokstäver som skiljer de båda varelserna åt.
E. menar att designförespråkare har fel på två punkter: dels eftersom de utgår från att fixeringen av mutationerna måste ske i en speciell sekvens, att de fixeras en efter en (s 140). Och dels för att de glömmer att det handlar om sexuell fortplantning med rekombination vilket gör att fixeringen går mycket fortare (s 141).
På den första punkten har E. fel. Haldanes dilemma förutsätter inte någon strikt sekventiell fixering, och ingen ID-förespråkare har heller någonsin hävdat det. För alla rimliga parametrar kommer väntetiderna för koordinerade mutationer (dvs. mutationer som måste inträffa tillsammans för att ha ett selektionsvärde) att vara fullständigt orealistiska (av storleksordningen miljarder år) alldeles oavsett ordningen på dem. Sant är det däremot att väntetidsproblemet blir ännu värre när mutationer dessutom måste inträffa i en specifik sekvens.
Detsamma gäller den andra punkten under realistiska förhållanden. En studie från 1998108 visade att
rekombination minskar väntetiden tills en ny genotypisk kombination först uppträder, men effekten är liten jämfört med mutationstakten och populationsstorleken.
Bara under mycket speciella omständigheter kan rekombination visas ha någon större inverkan på fixeringshastigheten, och det är när flera fördelaktiga mutationer uppstår oberoende av varandra i en population och konkurrerar med varandra, så kallad ”klonal interferens”.109 Detta kan möjligen vara relevant för bakterier där oddsen för uppkomsten av fördelaktiga (men inte konstruktiva – se kap 8 ovan) mutationer är förhållandevis hög, men knappast för mer komplexa organismer som människor. Väntetidsproblemet tillämpat inom området människans ursprung ger dystra resultat för de darwinistiska mekanismerna.110
E. anser att väntetidsproblemet är löst eftersom en datorsimulering från 2019 visar det. (s 141) Två andra undersökningar111 visar på motsatsen. Det minsta man kan säga är att man alltid bör förhålla sig skeptisk till datorsimuleringar, i synnerhet när de gör anspråk på att ge resultat som motsäger det vi intuitivt inser, som det inledande hårtussexemplet. Man kan då misstänka en bakomliggande bias. Faktiska observationer är viktigare.
Han nämner två sådana exempel. Dels med colibakterier (s 139) och dels den lilla växten backtrav (s 141).
Min hypotes
Colibakterierna utvecklade resistens mot antibiotika genom att fixera fyra samverkande mutationer i populationen inom loppet av tio timmar. Jag hade inte läst den här rapporten, men utifrån vad jag tidigare behandlat (s 48-49) drog jag slutsatsen att det här handlar om en enstaka punktmutation. När en viss fördelaktig punktmutation väl ägt rum, vilket är högst förväntat i en bakteriepopulation på miljarder individer, kan selektionen starta. Selektionskoefficienten är mycket hög eftersom individer utan mutationen sannolikt dör eller inte kan föröka sig.
Facit
Jag sökte sedan upp artikeln. Det visade sig stämma precis. (Jag ber icke-naturvetare om ursäkt för den här lilla djupdykningen). Antibiotikans (ciprofloxacin) primära mål är enzymet DNA-gyras, som har en viktig funktion i att reglera DNA-molekylens struktur genom att lösa upp överspiralisering och trassel som kan uppstå under replikation och transkription. Genetiska analyser har visat att resistens mot denna klass av antibiotika ofta kan uppstå till följd av punktmutationer i gener som kodar för DNA-gyras.112
I en normal bakteriecell binder antibiotikamolekylen till DNA-gyraset, vilket leder till att enzymets funktion hämmas och DNA-skador uppstår. Detta förhindrar celldelning och leder till att bakterien dör. En mutation kan medföra att enzymets bindningsställe förändras, så att antibiotikamolekylen inte längre binder lika effektivt. Därmed minskar den hämmande effekten, vilket ger bakterien ett selektivt övertag.
Ytterligare punktmutationer kan därefter uppkomma, och vissa av dessa kan försämra antibiotikans bindning ytterligare. Dessa mutationer gynnas av naturligt urval och sprids i populationen. Inom loppet av timmar kan därför fullt resistenta bakterier dominera populationen och fixeras, tack vare bakteriens korta generationstid på cirka 20 minuter.
Hur kunde jag gissa så här rätt? Tack vare att jag mindes Michal Behes forskningsresultat (se kap 15 ovan). När ”avståndet” till en selektiv fördel är en enstaka mutation är det en enkel match när populationerna är oerhört stora. När en egenskap väl har uppkommit kan mutationer och selektion snabbt finslipa systemet.
Men är detta då ett exempel på att en ny egenskap har uppstått? Nej. Den fördelaktiga mutationen är i vanlig ordning destruktiv till sin karaktär (se kap 8) eftersom den minskar specificiteten i en kemisk bindning. Minskad specificitet är alltid kopplat till minskat informationsinnehåll.113
Backtrav
Slutligen E.s backtrav, Arabidopsis thaliana. På sidan 141 argumenterar han så här:
Plantorna utsattes för stress och det visade sig att en oväntat stor del, upp till 95 procent, av dem hade selekterats bort i en generation. Ändå kunde de fruktsamma plantorna i experimentet täcka förlusterna. Det betyder att åtminstone i det här fallet måste Haldanes siffra om 300 generationer till fixering vara överdriven – det kan uppenbarligen gå betydligt snabbare.
Här blandar E. ihop korten i flera avseenden.
Han anger ingen referens, så jag blev tvungen att ge mig ut på nätet och söka. Det visade sig att det inte finns någon forskning på Arabidopsis thaliana med de slutsatser som han gör gällande. Av allt att döma har han förväxlat Arabidopsis thaliana med Arabidopsis suecica. Men den senare är en hybrid mellan de båda arterna Arabidopsis thaliana och Arabidopsis arenosa. Och som sådan är den polyploid, det vill säga den har en dubbel genuppsättning som gör att varje individ bär en mycket större andel av hela populationens genvarianter än vad som är normalt. Den här inbyggda redundansen är anledningen till att en så stor andel av genvarianterna inte försvann med de 95 procenten som aldrig fick sätta frö.
Bortsett från detta har exemplet väldigt lite bäring på Haldanes dilemma som handlar om hur lång tid det tar för gynnsamma mutationer att spridas via långa perioder av upprepad selektion. Men här har vi i stället en variation som redan existerar förpackad i några få individer och utan någon lång selektionskedja. Så faktiskt ett dubbelfel av E.. Väntetidsproblemet står kvar. Som ett argument mot evolution precis som E. konstaterar på kapitlets sista rad, men samtidigt indirekt för design.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 22. Det elfte argumentet: Universums finavstämning
Här finns inte utrymme för någon noggrann genomgång av finavstämningsargumentet (s 143), men är du intresserad kan du själv läsa om det på nätet. Sök då på ”universums finjustering” eller ”finetuning of the universe” som är vanligare beteckningar. Alternativt kan man söka på ”den antropiska principen”. Den förekommer, som E. mycket riktigt noterar, i två versioner. Den ena, den svaga varianten, säger att intelligent liv bara är möjligt i ett finjusterat universum så det är 100% förväntat att vårt universum är det. Alltså finns inget som behöver förklaras. Den starka varianten är att det finns en bakomliggande avsikt med att universum är finjusterat.
I korthet handlar det om att naturens grundläggande lagar och konstanter verkar vara exakt inställda för att möjliggöra liv. Om dessa värden hade varit bara marginellt annorlunda, skulle stjärnor, atomer eller kemiska processer som krävs för liv inte kunna existera.
Det här förhållandet är naturligtvis hand i handske ett designperspektiv där man tänker sig en värld skapad av Gud. Däremot är det inte alls förväntat ur ett naturalistiskt perspektiv där det i stället är blinda och likgiltiga naturkrafter som antas vara orsaken. Med en sådan världsbild behöver man bort… förlåt … förklara illusionen av att Någon finjusterat alltsammans. Det är E.s avsikt med det här kapitlet. Hur resonerar han då?
Han börjar med att konstatera att det finns något missvisande med begreppet finavstämning som sådant, eftersom det skulle kunna misstolkas som att det måste finnas en intelligent orsak som gjort avstämningen. Det är förstås lika möjligt att en sådan tolkning är en logisk konsekvens av de observationer man gör, men den tanken är E. inte bekväm med. 114
Därefter skriver han att ett finavstämt universum snarare är ett argument för evolution:
Då behövs ingen designer för att förklara livet. Det är snarare förekomsten av liv i ett livsfientligt universum som skulle kräva en designer. (s 144)
Det är förvisso sant att det är oerhört trixigt att bygga ett hus i en tät granskog. Det är lika sant att det är betydligt lättare att bygga huset om man först sågar ner träden och bereder marken. Men frågan är om det skulle vara ett hållbart argument för att huset kan uppstå utan närvaro av några intelligenta aktörer som arkitekt och snickare. Själv tycker jag inte det.
Sedan lägger E. fram sex argument:
1. Det finns så många andra planeter i andra solsystem än vårt (exoplaneter) i universum, så det är inte alls säkert att jorden är så speciell (146).
2. Det skulle kunna vara så, att naturkonstanterna som förefaller så fininställda kanske måste ha just de värden som de har på grund av någon idag okänd naturlag (s 147).
3. Naturkonstanterna kanske skulle kunna vara mycket bättre finjusterade än de är, till exempel i en annan dimension enligt modern strängteori (s 149).
4. Strängteorin möjliggör förekomsten av oändligt många universum. Denna syn är inte lösa spekulationer, utan grundar sig på ”känd fysik” och ”strikt matematik” till skillnad från designteorin som
är ren spekulation. Den grundar sig inte i någonting; designern är helt gripen ur luften. (s 149)
5. Det går inte att räkna på saker man inte har siffror på och därför kan man inte ha finavstämning som designargument. (s 149)
6. Finavstämningsargumentet är inte falsifierbart (s 150).
Några korta kommentarer i tur och ordning:
1. Ja, man tror sig i nuläget ha evidens för ungefär 6000 exoplaneter. Av dessa tror forskare att två till fyra av dem (0,03-0,07 procent) verkar ”lovande” i bemärkelsen att de kanske har en fast yta (stenplanet) och någorlunda stabil bana runt en någorlunda lugn stjärna. Något vatten eller atmosfär har ännu inte påvisats på någon av dem. Så om man föredrar evidens framför spekulationer så tycks jorden tämligen speciell.
Det talas ibland om att göra en höna av en fjäder. Ibland ger nyhetsrubrikerna intrycket att om det möjligen finns vatten på en planet så finns där nog liv. Att göra en höna av en fjäder är en förhållande enkel match i jämförelse. Fjädern innehåller inte bara DNA och proteiner utan all information som kodar för en höna. Vatten innehåller ingetdera.
2. Om det finns en naturlag som bestämmer att naturkonstanterna måste vara så livsbejakande som de faktiskt är, så förskjuter man bara problemet ett snäpp bakåt-uppåt. Hur uppstod den där finurliga lagen?
3-4. Enligt strängteorin kan det finnas 10500 universum i andra dimensioner än vårt eget. E. har helt rätt i att man använda matematik för att räkna på sådana, men det gör dem knappast mindre spekulativa för det. Vi kommer aldrig att kunna nå de här andra dimensionerna med våra instrument därför att de per definition saknar kontakt med vår värld. Inga observationer kräver, och inga experiment vare sig bekräftar eller falsifierar strängteorin. Att avfärda design genom att hänvisa till hypotetiska multiversum är därför inte mer relevant än det låter.
5. E. har också rätt i att det inom forskarvärlden råder delade meningar om hur stora frihetsgrader det finns hos de naturkonstanter som gör vår värld möjlig och beboelig. Men det är inte detsamma som att universum skulle sakna finavstämning. Debatten handlar främst om till vilken grad. Det råder praktiskt taget total konsensus bland forskare om att naturens grundläggande konstanter verkligen ligger inom mycket snäva intervall som tillåter materia, stjärnor och liv att existera. Det man heller inte är eniga om är varför det förhåller sig på det viset.
En analys som gjordes av filosofen Roberto Fumagalli förra året115 stöder designhypotesen. Han menar att sannolikheten att finjustering uppstår genom slumpfaktorer är avsevärt lägre än att den uppkommer genom icke-slumpfaktorer. Och det faktum att ”det finns ett tryck på vetenskapliga eller filosofiska teorier att undvika högst osannolika sammanträffanden” talar till förmån för icke-slump-förklaringen (design). Observera att Fumagalli inte själv tar någon personlig ställning i frågan.
6. Det stämmer att finavstämningsargumentet inte är falsifierbart utifrån dagens kunskapsläge, men som vi sett gäller detsamma för strängteorin med dess multiversum.
Finavstämning – slutsatser
För att sammanfatta det sagda kan jag hålla med E. om att finavstämning inte kan användas som ”bevis” för design. Men det går heller inte att avfärda det utan att på samma grunder avfärda eventuella alternativ. Däremot har vi sett att det finns vetenskapligt stöd för att design är en troligare förklaring än slumpfaktorer till att universum har de egenskaper det har.
Jag vill också tillägga att jag tycker att E. har en poäng även i det han skriver på sidan 146. Han invänder där mot de antropocentriska116 uttalandena i Gonzales och Richards The Privileged Planet:
Kanske det om några hundra miljoner år uppträder en annan form av intelligent liv på jorden, som kommer att vara det liv som vår planet egentligen är inrättad för? På tal om tid kan man också fråga sig varför designern behövde bortåt 14 miljarder år för att åstadkomma oss. (s 146)
Som kreationister hävdar vi att det finns fog för en antropocentrisk syn i vissa avseenden, eftersom människan skapades till Guds egen avbild med uppdraget att bruka och förvalta jorden. Men vi tar avstånd från ID-rörelsen åldersperspektiv. Det skapar ibland onödiga problem, precis som det E. påtalar.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 23. Det tolfte argumentet: Maskinargumentet
Bland det mest fascinerande som framkommit inom molekylärbiologin det senaste halvseklet är upptäckten att celler innehåller komplexa proteinmaskiner. De här strukturerna består av komplex (grupper) av mindre proteinmolekyler med specifik tredimensionell struktur. Sekvensen (den inbördes ordningen) av deras aminosyror (se kap 16 ovan) ger dem egenskaper som bland annat bestämmer hur de fäster till varandra så att hela komplexet får en viss bestämd funktion i cellen.
Exempel på sådana maskiner är enzymet DNA-polymeras, som möjliggör kopiering och korrekturläsning av DNA i samband med celldelningen. Ribosomen tillverkar proteiner med hjälp av instruktionerna i RNA. Enzymet ATP-syntas tillverkar ATP som är alla cellers gemensamma energivaluta. Kinesiner transporterar behållare av byggnadsmaterial till olika delar av cellen där det finns behov genom att helt sonika traska med små myrsteg (gigantisk överdrift). Och spliceosomen hanterar så kallad RNA-splitsning – den fascinerande process genom vilken en enda RNA-sträng kan klippas och klistras ihop till tusentals olika varianter som i sin tur översätts till lika många funktionella proteiner i ribosomerna.117
Och, förstås, den numera klassiska bakterieflagellen. Denna elektriska motor som återfinns i bakteriers cellväggar består av ett 40-tal proteinenheter som sätter flagellen (”propellern”) i rotation med en hastighet av upp till 100 000 varv/min och kan slå om till backläge inom loppet av ett kvarts varv. Den är utrustad med ett effektivt bromssystem, och dessutom med en växellåda som gör att flagellen kan frikopplas vid behov. Och därtill är den integrerad i ett reglersystem som gör det möjligt för bakterien att justera sin transportriktning efter skillnader i ljusstyrka och kemiska gradienter. Och allt med en verkningsgrad som får vår tids konstruktörer av elektriska motorer att rodna av avund. Faktum är att E. har en bild på flagellmotorn i boken, men inte under maskinkapitlet utan i det som behandlade Förklaringsfiltret (fig 7 s 121).
Det är viktigt att ha den här kunskapsbakgrunden innan man tar del av E.s vederläggning av ID:s så kallade maskinargument. Först då är det möjligt att värdera hans resonemang. Låt oss först se vilka hans motargument är:
1. E. anser det felaktigt att kalla cellens olika proteinkomplex för maskiner eftersom de verkar på en annan storleksskala där viskositeter118 och molekylrörelser skulle ställa till det för en vanlig maskin. Därför är analogin falsk. (s 152-154)
2. [D]et är en metafor man bör undvika. (s 154)
3. Cellernas proteinkomplex är självmonterande, och dessutom bryts de ständigt ner och återmonteras. Det gäller inte de maskiner som människor konstruerar (s 154).
4. E.s laptop har inte designats av enda designer, utan är ett resultat av en lång teknologisk utvecklingsprocess som inbegriper tusentals forskare och ingenjörer under hundratals år, det vill säga en evolutionär designprocess genom selektion av de bästa idéerna (155).
5. Om en designer skapat molekylära maskiner så skulle vi förvänta oss att vederbörande
kunde skapa unika, fulländade konstruktioner, utan spår av någon evolutionär designprocess. Vad vi inte skulle förvänta oss vore att se spåren av en designer som plockar och donar med samma delar. (s 156)
Låt oss nu i sedvanlig ordning behandla dem ett i taget för att pröva om de håller:
1. Antag att en nanoingenjör bestämmer sig för att som doktorsavhandling konstruera en maskin som är tänkt att fungera inuti en levande cell. Hans eller hennes första uppgift blir att undersöka de grundläggande funktionsprinciperna för den miljö där maskinen ska arbeta. Konstruktionen utformas sedan så att den kan fungera på just denna nanoskala där värmerörelser och reynoldstal119 är extremt annorlunda än de vanligtvis är. Vore det inte ganska märkligt om ingenjören efter slutfört projekt möts av beskedet att avhandlingen inte kan godkännas eftersom maskinerna är för små för att få lov att heta så? Jag tycker det. Det finns ingen rationell anledning att koppla definitionen av maskin till dess storleksordning. Det skulle aldrig ske om det inte vore föranlett av en agenda.
2. Cellens nätverk av samarbetande molekylära maskiner möjliggör livets funktioner. Själva uttrycket ”molekylär maskin” har länge varit både erkänt och använt inom molekylärbiologin. Om du som läser detta betvivlar det så föreslår jag att du gör en sökning med ”molecular machine” på artikelbaser som till exempel PubMed eller Google Scholar. Ett par exempel: The ATP Synthase — a Splendid Molecular Machine från 1997 och The 26S Proteasome: A Molecular Machine Designed for Controlled Proteolysis från 1999.
Kemister inom området organisk syntes har numera lyckats tillverka egna så kallade nanomaskiner, även om deras kreationer är oerhört mycket blygsammare än de naturliga motsvarigheterna. I samband med Nobelpriset i kemi 2016 meddelade Kungliga Vetenskapsakademien:
Nobelpriset i kemi 2016 tilldelas Jean-Pierre Sauvage, Sir J. Fraser Stoddart och Bernard L. Feringa för deras design och produktion av molekylära maskiner. De har utvecklat molekyler med kontrollerbara rörelser, som kan utföra en uppgift när energi tillförs.
Begreppet molekylär maskin förefaller alltså både passande och okontroversiellt, och enligt många ordboksdefinitioner utgörs maskiner av ”kombinationer av sammankopplade komponenter som är arrangerade för att överföra eller modifiera kraft med syftet att utföra nyttigt arbete”, alltså i linje med Vetenskapsakademiens formulering.120
E. menar alltså att det inte är berättigat att använda sig av en analogi mellan de här proteinkomplexen och nanomaskiner därför att det finns skillnader mellan dem, som till exempel i deras funktionsprinciper. Men om de båda hade varit oskiljaktiga så skulle det givetvis inte råda någon analogi över huvud taget mellan dem; då skulle det ju vara två identiska företeelser. Hela analogibegreppet bygger på just de olikheter som gör att nanomaskinen och proteinkomplexet verkligen går att särskilja.
Sedan kan man förstås tillägga att den främsta skillnaden mellan mänskligt konstruerade maskiner och naturens egna är att de senare som regel är vida överlägsna de mänskliga motsvarigheterna när det gäller såväl sinnrikhet, driftsäkerhet, verkningsgrad som annat. Jag har diskuterat det här tidigare med E., och den som vill ta del av en utförligare redogörelse kan göra det via de båda länkarna.121, 122
3. Ja, det stämmer verkligen att livets molekylära maskiner är både självmonterande och dynamiska, och jag har skrivit om det i andra sammanhang.123, 124 Men frågan är om detta faktum är ett argument som talar emot design – eller kanske precis tvärt om.
Att en molekylär maskin kan vara självorganiserande och montera sig själv beror på att de enskilda proteinerna i komplexet är försedda med plus- och minusladdningar fördelade på ett sådant sätt att de bara passar ihop på ett enda sätt.
För några år sedan konstruerad Caroline Larsson vid Linköpings universitet ett självmonterande modellvirus125 genom att fästa små magneter med nord- och sydpol på genomtänkta ställen på små plastbitar. Man lägger bitarna i en påse och skakar runt en stund och vips så bildas små virusbollar. Hennes syfte, precis som E.s i den här boken, är att gymnasieelever ska inse att naturens molekyler har självorganiserande egenskaper och att det vi intuitivt uppfattar som design i naturen kan vara något skenbart. Jovars, men hur fick naturen den egenskapen, månntro? Själv fick hon ägna många timmars tankemöda och tålamod för att få till sina modeller.
Caroline skrev: ”Det är viktigt att man tillhandahåller trovärdiga förklaringar”. Visst är det så. Laborationen är mycket illustrativ för att visa att små delar spontant kan självorganisera sig till en komplex struktur, men givetvis bara under villkoret att de dessförinnan är noggrant genomtänkta och förberedda av en intelligent designer, i det här fallet Caroline. Laborationens verkliga implikation – att självmonterade virus vittnar om en annan Designer – Gud – får man förstås inte berätta för eleverna ifall Skolinspektionen skulle komma på besök. Man hoppas att eleverna inser det ändå, men det är tyvärr långtifrån säkert. Det är därför det behövs bibeltroende lärare som prenumererar på magasinet Genesis.
Dit jag vill komma är att det krävs avsevärt mycket mer och högre organiserad information för att koda för en självorganiserande struktur i DNA än en som inte är det. Och analogt tog det givetvis Caroline mer tankemöda att klura ut hur hon skulle placera ut sina magneter på lämpliga ställen i förhållande till om hon inte behövt göra det.
4. E. jämför den teknologiska utvecklingen av hans laptop med hur evolutionen har utvecklat molekylära maskiner genom selektion. Jag tycker att det är lika häpnadsväckande varje gång evolutionister presenterar argument som detta. De visar därmed att de inte kan se skillnad mellan en aktiv teleologisk (målstyrd) intelligent designprocess (som att designa en dator) från en passiv planlös ointelligent process via slumpmässiga mutationer.
Skillnaden mellan dem är monumental: den ena sker genom input av väldiga mängder meningsfull funktionell information medan den andra saknar det. Det är det som är orsaken till att naturligt urval bara kan modifiera sådant som redan existerar.
E. säger själv det när han citerar ”Galls lag” (s 155):
Ett komplext system som fungerar har alltid evolverat från ett enkelt system som fungerar. Det omvända påståendet tycks också vara sant: Ett komplext system som designas från första början fungerar aldrig och kan inte fås att fungera. Man måste börja om från början med ett fungerande enkelt system.
Galls andra mening säger att om ofullkomliga människor från början detaljplanerar och konstruerar ett hus, webbplats eller organisation så kommer man garanterat att få problem med tid- och resursslukande omorganisationer. Det är därför bättre, menar han, om man börjar i liten skala och är flexibel i sin designprocess. Märk väl att Galls lag alltså handlar om en mänsklig intelligent designprocess och inte någon slumpbaserad sådan. Från ID:s perspektiv kan vi naturligtvis invända att när en fullkomlig Gud skapade livets system, så var Han fullt kapabel att skapa optimala system redan från början.
Principen fungerar när den tillämpas på företeelser som IT och administration men dåligt på arkitektur och ingenjörskonst. Anledningen är att hierarkisk struktur inte kan skapas genom att bara addera moduler bottom-up. Det blir bara just en samling moduler. Hierarkiskt uppbyggda föremål som ett hus, en bil eller ett datorprogram kan bara skapas top-down, det vill säga genom att först skapa en grundstruktur (de stora dragen i ett byggprojekt och dess logistik med rör- och ledningsdragningar med mera) och via förändringar på lägre hierarkiska nivåer (exakt var fönstren ska sitta) till de lägsta hierarkiska nivåerna (färgen på tapeterna).126
Det är just därför som Galls lag inte är tillämpbar på uppbyggnaden av biologiska strukturer som bevisligen är hierarkiskt arrangerade. Den enda hypotetiska tillämpningen på biologin är om man som E. utgår från att evolutionen har uppstått i en historisk sekvens från det enkla till det komplexa (från en bacill till en människa). Bara med
5. Här spekulerar E. om hur Skaparen borde eller inte borde ha gjort när han skapade livets strukturer. Kanske har han glömt att han nyss anklagat ID-anhängare för att spekulera om designerns egenskaper?
E. skriver som sagt att vi inte borde förvänta oss
att se spåren av en designer som plockar och donar med samma delar. (156)
Men det där stämmer inte. När vi ser just den sortens spår i naturen så bekräftar det snarare än motsäger ID-perspektivet, även om det inte varit någon uttalad prediktion.
Det förefaller nämligen som om det var den principen Skaparen arbetade efter när han designade livet i alla dess former. Han programmerade modulärt, precis som man gör inom modern programmeringsteknik. Ingen duktig programmerare bygger idag sina datorprogram ”från scratch”. I stället går vederbörande ut på internet och laddar ner färdigskapade moduler för de funktioner han eller hon vill ha i sitt program. Det kan handla om hur data matas in, hur de lagras eller hur de matas ut (till exempel till en bildskärm). Sådana moduler förekommer i princip i alla mjukvaruprogram.
På liknande sätt finns det genetiska programmoduler för en stor mängd funktioner i olika slag av levande organismer utan att evolutionsbiologer menar att de har sitt ursprung i någon gemensam urmoder. Sådana här genetiska moduler är legio och brukar med ett finare namn kallas molekylära parallellismer. De brukar förklaras av så kallad konvergent evolution, vilket egentligen inte är någon förklaring utan snarare en etikett.127 Innan Google tog bort funktionen vet jag att jag fick hundratusentals träffar på sökorden ”convergent evolution” vilket ger en antydan om hur utbrett fenomenet är i den levande världen.
Slutligen använder E. biologisk evidens till att argumentera mot design, men kritiserar samtidigt (s 155-156) Anders Gärdeborn för att använda biologisk evidens som argument för design. Man borde med all rätt fråga sig varför det ena skulle vara ok men inte det andra?
Det här betyder att ingen av E.s fem invändningar mot maskinargumentet håller för en kritisk granskning.
↩ Tillbaka till slutet av introduktionen
![]()
Kapitel 24. Det trettonde argumentet: Det intuitiva argumentet

Det krävs inte särskilt mycket för att vi intuitivt inser att någonting gjorts med avsikt. Ett par stenar placerade på varandra längs stigen, ett par bokstäver i sanden eller en bit metalltråd på marken. I samtliga fall använder vi vår kunskap och livserfarenhet till att dra den slutsatsen.
Vi har förmågan att detektera ännu subtilare saker, som till exempel om någon säger något men inte uppriktigt menar det, eller om det är en AI-assistent som skrivit ett textavsnitt eller en levande människa (det så kallade Turing-testet).
Det här beror på att vi är utrustade med system som hämtar information från omgivningen (input), bearbetar den medvetet och omedvetet (kontrollcentrum, algoritmer) och som sedan levererar (via någon form av effektor) ett resultat i form av en ändamålsenlig handling eller som i det här fallet en slutsats (output).128
Det låter tekniskt, men är bara teknikers sätt att beskriva en process som vi spontant använder oss av varje stund av våra liv utan att ens reflektera över det, det lilla barnet såväl som den vuxne. Och det gör också alla andra levande varelser, men inte i samma grad.

Stenstatyer på Påskön. Foto: Bjørn Christian Tørrissen derivatverk: George Tsiagalakis, CC BY-SA 3.0 Wikimedia Commons.
Men låt oss gå vidare till saker som inte är lika subtila. De berömda stenstatyerna på Påskön i sydöstra Stilla havet bär omistliga kännetecken på design. Utan att vi har någon metod för att kvantifiera dessa kännetecken kan vi konstatera att de är formade på ett både komplext och specifikt sätt så att vi omedelbart och intuitivt drar slutsatsen att de föreställer människor.
Som vi konstaterat tidigare menar E. att det handlar om falska analogier när vi tillämpar samma resonemang på den levande världen (s 6, 34, 53, 78). Vi vet ju av erfarenhet att människor skapar former i sten, men vi har aldrig observerat Gud skapa en levande varelse och följaktligen är analogin irrelevant.
I fallet med levande varelser förordar E. ointelligent design, vilket i praktiken är en självmotsägelse eftersom ordet design förutsätter en intelligent designer. Han är givetvis långtifrån ensam om detta. Illusionen om en design utan designer är utbredd bland ateistiska och agnostiska naturvetare (och många andra). Tydligast kommer det till uttryck hos personer som Francis Crick129 och Richard Dawkins.
Dawkins:
Levande materia … förmedlar en elegant illusion av avsiktlig design.130Biologi är studiet av komplicerade företeelser som ger sken av att ha blivit designade för ett syfte.131
Crick:
Biologer måste ständigt ha i åtanke att vad de ser inte har blivit designat.132
Man kan alltså konstatera att designintuitionen är djupt rotad i den mänskliga naturen.133 Så djupt att inte ens fullfjädrade ateister som de båda citerade herrarna kan undvika att ge uttryck för den. Men inte genom något erkännande av den utan i ett aktivt avståndstagande. Designintuitionen förefaller dessutom nedärvd, eftersom även små barn har den.134
I sin bok Undeniable visar proteinforskaren Douglas Axe på hur vår intuition är dessutom har en djup vetenskaplig förankring eftersom den i grunden baseras på matematisk statistik. Jag har skrivit om det i andra sammanhang.135
Men E. håller naturligtvis inte med om det jag just skrivit. Han erkänner i inledningsmeningen till sin vederläggning av det här trettonde argumentet att
Det här är en verklig tungviktare. Det är det centrala argument som funnits mer eller mindre uttalat i bakgrunden under hela diskussionen, själva urargumentet: att man känner igen design när man ser den. (s 157)
Han har helt rätt. Designintuitionen är den egentliga kärnan i ID. En biblisk kreationist skulle omedelbart referera till Romarbrevets första kapitel:
Det man kan veta om Gud är uppenbart bland [människor], Gud har ju uppenbarat det för dem. Ända från världens skapelse ses och uppfattas hans osynliga egenskaper, hans eviga makt och gudomliga natur genom de verk som han har skapat. Därför är de utan ursäkt. (Rom 1:20)
Man skulle kunna säga att ID:s roll är att sätta strålkastarljuset på det som teologer brukar kalla ”den allmänna Gudsuppenbarelsen”, att Gud gjort sig fullt synlig för människan genom sin skapelse. Det är också denna strålkastare som E. upplever sig kallad att släcka ner. Varför? För att han anar en politisk och religiös agenda i bakgrunden. Vi återkommer snart till det.
Men först ska vi se vilka argument E. presenterar för att ta udden av det här mest centrala ID-argumentet. Han återkommer i inledningen av det här kapitlet till proteinforskaren Douglas Axes 13-åriga studie vid Cambridgeuniversitetet. Han upprepar sin tes att Axe inte tagit hänsyn till att proteiner kan evolvera stegvis (s 158). Jag har redan analyserat hans skäl till det. Han fortsätter sedan med att lägga fram fyra argument till varför intuitionsargumentet inte skulle vara giltigt:
1. Designintuitionen är enligt psykologer
en rest av ett naivt och outvecklat tänkande. (s 159)
2. Att tillämpa designintuitionen på biologin bidrar till vetenskapsförakt och att fjärma allmänheten från forskarvärlden. (s 160-161)
3. Intuitionen kan bedra oss. Det finns saker som är sanna fast de är anti-intuitiva (som relativitetsteori, kvantmekanik och – menar han – evolution). (s 162)
4. Intuitionsargumentet handlar inte om sakfrågor, det bygger uteslutande på känslor. (164)
Låt oss se:
1. E.s åsikt att människans designintuition är någon sorts rudiment (kvarleva) av ett primitivt sätt att tänka förutsätter att evolutionen faktiskt har ägt rum. Men man kan inte argumentera mot design genom att utgå från evolutionens faktum utan att det blir cirkelresonemang. Därför är detta varken mer eller mindre än E.s personliga tyckande som kommer till uttryck. Ingenting hindrar honom från att tänka så, men för den läsare som frestas att tänka i samma banor är detta viktigt att veta om.
2. Vetenskapsförakt gror knappast i en miljö där argument öppet förs fram och diskuteras på ett sakligt sätt och i en konstruktiv anda. Annorlunda är situationen när fakta undanhålls, kritik avfärdas reflexmässigt och spekulationer anses trovärdigare än faktisk evidens och presenteras med tvärsäkerhet. Då talar vi inte längre om vetenskaplighet utan om dogmatism. Sådan är inte värd allmänhetens respekt.
3. Och intuitionen då, som är grundbulten i diskussionen? Ja, eftersom E. inte kan avfärda intuitionen som vare sig irrelevant eller relevant, men ändå måste ”oskadliggöra den”, så enda möjligheten är att beröva den dess relevans i just designsammanhanget. Han skriver därför:
Slutsatsen av allt detta blir att man lugnt kan förlita sig på sin intuition och sitt sunda förnuft – utom när man inte kan det. (s 163)
Det där sista är förstås lika klargörande som det låter, men det sammanfattar ganska väl E.s budskap: när vi som vanliga medborgare ska ta ställning i frågan om skapelse-evolution så måste vi lägga vår intuition och vårt sunda förnuft på hyllan, annars kommer vi att hamna fel. Bara det bör vara en tydlig indikation på att det finns en hund begraven.
4. Till sist det här med känslor. Det är precis känslor och subjektivitet som E. appellerar till med sina båda avslutande bilder på en sten och en katt (s 164). Han vill illustrera sin tes att designteoretiker bygger sina argument på en känslobaserad intuition.
***
Jag finner det anmärkningsvärt att E., som vill framstå som expert på intelligent-designrörelsen, totalt missar ID:s poäng. Det designteoretiker som Dembski, Meyer och Axe ständigt arbetar med är att utveckla modeller för designdetektering som utesluter både falska positiver och falska negativer så att de bara ”fångar in” fall där design är reell. Falska positiver illustreras av E.s båda bilder – varken stenens eller kattens mönster är resultat av design, utan är slumpens verk.136
Falska negativer är klurigare. Man kan aldrig vara säker på att någonting som ser slumpmässigt ut verkligen är det. Man kan till exempel skriva en slumpmässig text eller sprida ut föremål på ett sätt som ser ut att vara ett resultat av tillfälligheter men som inte är det. Att avslöja sådant kan vara avgörande inom forensisk vetenskap när en kriminell medvetet försökt utplåna spåren efter sin brottsliga aktivitet.
Det kan också ha relevans i biologiska sammanhang. Fördelaktiga mutationer kan ibland misstas för att vara slumpmässiga, men i praktiken vara riktade åt ett ändamålsenligt håll därför att organismen är utrustad med genetiska system för att åstadkomma just detta.137
I båda dessa fall är emellertid designdetekteringsmodellernas tillkortakommanden ”åt rätt håll”, det vill säga de utesluter design aningen oftare än de borde, för säkerhets skull.
Ska vi alltså lyda E.s råd och sluta använda oss av vår intuition och vårt sunda förnuft när det gäller en av de viktigaste frågorna i tillvaron – den varifrån vi kommer? Absolut inte.
Jag har nu granskat E.s tretton argument mot intelligent design. Håller de? Det är upp till dig som läsare att bedöma efter att nu ha fått del av båda sidorna av myntet.
↩ Tillbaka till innehållsförteckningen
![]()
AVDELNING III Rörelsen för intelligent design
Bokens sista och avslutande del behandlar ID som rörelse.
Kapitel 25. ID som vetenskapligt alternativ
I det här kapitlet sammanfattar E. de slutsatser han kommit fram till i bokens två första avdelningar. Eftersom innehållet är helt redundant misstänker jag att hans förhoppning är att detta kapitel ska komma att användas som ett litet sammanfattande kompendium i utbildningen på gymnasier och högskolor.
Han repeterar här sina uppfattningar om de tretton ID-argumentens påstådda tillkortakommanden (s 167-168), hur ovetenskaplig ID är därför att den enligt honom inte går att falsifiera (s 168-170) och för att den inför en odefinierad designer (s 177), att det inte finns några vetenskapliga publikationer av företrädare för ID (s 170-174) och att ID är pseudovetenskap (s 178-179).
Katt bland hermelinerna
Som en liten katt bland hermelinerna har han sedan ett avsnitt om den kontroversiella svepeduken i Turin. Av vilken anledning vet jag inte, eftersom jag aldrig under alla år sett den behandlas i vare sig ID-litteraturen eller i kreationistisk litteratur, utom möjligen i ytterst skeptiska ordalag. Jag kan bara tänka mig en orsak och det är ”skuld genom associering”, ett av de klassiska logiska felsluten. Jag återkommer till det där i slutet av dokumentet.E. avslutar kapitlet med formuleringen:
Så om man nu är klar över att ID inte kan tävla med naturvetenskapen, varför dessa attacker på etablerad vetenskap? (s 181)
Du som läst boken och håller med honom om att ID attackerar etablerad vetenskap får mer än gärna återkoppla till mig och precisera på vilket sätt. Jag hävdar nämligen bestämt att det enda sättet att komma till den slutsatsen är om man sätter likhetstecken mellan etablerad vetenskap och evolutionistiskt historieberättande.
Paradoxalt nog inleder E. det här kapitlet med att illustrera precis det här. Han slår nämligen där ett slag för den kontroversiella hjälphypotes till evolutionen som har föreslagits av några forskare inom tredje vägens evolutionsbiologi, nämligen den om självorganisation. Jag har i andra sammanhang behandlat den här hypotesen138 och jag kan intyga att den är långt mer naturmagi än vetenskap.
En formulering som:
Man kunde exempelvis tänka sig att materien har självorganiserande egenskaper man inte känner till idag, och att detta är orsaken till att komplicerade system uppkommer. (s 167)
visar att E. utan att tveka är beredd att lämna beprövad vetenskap bakom sig och sträcka sig efter okända mekanismer med förmåga att skapa komplicerade system – så länge de inte har något att göra med den där Designern, vill säga.
E. fortsätter:
Och det finns faktiskt belägg för att ordnade molekylära strukturer kan uppkomma spontant under vissa omständigheter. (s 167)
Utan tvekan! Men ordnade molekylära strukturer kännetecknar livlösa ting som sockerkristaller och markens mineraler. Spontana kemiska och fysikaliska processer åstadkommer ständigt sådant. Men det är inte detta som kännetecknar livets molekyler, utan i stället den icke-ordnade villkorlighet (se kap 18 s 67 om förklaringsfiltret) som karakteriserar sekvenserna i DNA. Om man ställer kaffekoppen på tangentbordet medan man går på toaletten har datorn själv skrivit 80 sidor med en högst ordnad text, alldeles av sig själv, när man kommer tillbaka. Det är hur en kaffekopp kan skriva en läsbar och meningsfull artikel som behöver förklaras!
E. refererar till en bok av Jeremy England: Every Life Is on Fire: How Thermodynamics Explains the Origins of Living Things. Det är ingen direkt anspråkslös boktitel, och den i sig borde vara tillräckligt för en person som gör anspråk på att företräda vetenskap att dra öronen åt sig. England är teoretisk fysiker, och sådana brukar ibland ha en tendens att missförstå kemi. Det gäller i högsta grad England, och ingenting tyder på att han har den minsta förståelse för förutsättningarna för hur livets molekyler skulle kunna bildas.
Det enda som kopplar Englands självorganisationshypotes till den materiella verkligheten är att liv följer termodynamikens lagar och att självorganisation kan kopplas till energitillförsel. Hans arbete är rent teoretiskt och bygger inte på några som helst observationer, varken i kemi eller biologi.
Därför är den enda reella empiriska grunden för E.s tankar kring spontan uppkomst av ”komplicerade system” i princip på samma nivå som att när solen skiner på en liten saltvattenspöl så bildas det med tiden spontant saltkristaller, vilket varje västkustbo noterat. Föreställningen att det med en sådan mekanism dessutom kan uppstå komplicerade eller komplexa ting rymmer också oförmågan att skilja mellan allmänt komplexa företeelser och specifikt komplexa sådana, vilket vi behandlade tidigare.
Det här kapitlet var därför ett lågvattenmärke. Återstoden av boken innehåller vissa poänger
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 26. Vad är intelligent design – egentligen?
E. inleder kapitlet med att ställa frågan: Om nu ID är ett så hopplöst misslyckat vetenskapligt projekt – varför fortsätter då den konservativa tankesmedjan Discovery Institute (DI) att pumpa in miljoner dollar i det? Lätt att svara på, menar han. Målet är att ersätta materialismen och dess destruktiva inflytande över samhälle och kultur och i stället införa teism, tron på Gud. Strategin för att nå det målet är att med hjälp av designteorin successivt påverka gräsrötterna, vanliga medborgare, till att skapa en politisk majoritet för en genomgripande samhällsomvandling. Det här, skriver E.
kan sammanfattas under namn av ”kristen nationalism” eller ”dominionism”. (s 184)
Dominionism, jag hade aldrig hört talas om ordet. E. förklarar:
Man utgår från övertygelsen att man har Guds kallelse att skapa ett kristet samhälle genom att ta kontroll över samhällets politiska och kulturella institutioner. (s 184)
Han fortsätter sedan med att beskriva nästa ism som jag inte heller hade hört talas om:
Den mest extrema grupperingen är nog kristna rekonstruktionister, som menar att samhället helt måste styras av kristna. (s 184)Rekonstruktionismen vill styra strikt efter Gamla testamentets lagar, vilket innebär att man vill se dödsstraff för homosexualitet, otrohet, hädelse, trolldom och en hel del annat. (185)
Nu har vi ID-rörelsens målbild klar, menar E.. Hur är det då med vägen dit?
Nu följer en historisk tillbakablick via den kända ”Scopesrättegången” i Dayton, Tennessee 1925, där kreationisterna förlorade slaget om skolan, men ändå ledde till en nedtoning av evolutionsinslagen i skolans läromedel. Sedan kom Sputnik-krisen som resulterade i 50-talets uppryckning av det amerikanska skolsystemet och en nysatsning på bland annat evolutionsteorin. Sedan poppade ett antal kreationistorganisationer upp som svampar ur jorden och Morris och Whitcombs klassiska bok om syndaflodsgeologi, The Genesis Flood såldes i hundratusentals exemplar. Kreationisternas hopp om att få ”creation science” på skolschemat grusades emellertid, först i Louisiana 1987 och sedan i Dover, Pennsylvania 2005.
Det är nu E. menar att ID kommer in på arenan, alltså som en sorts kamouflerad kreationism, uppbackad av DI:s program Center for Science and Culture med bland annat webbplatsen Evolution News & Science Today.139 Det är det nya vapnet för att angripa roten till problemet – den metodologiska naturalismen, fundamentet för den moderna vetenskapen. Slutligen rundar E. av resonemanget med att påminna om ID-rörelsens populistiska anti-intellektuella karaktär och hur man platsar bland
klimatskeptiker, vaccinationsmotståndare och plattjordsförespråkare. (s 193)
Det mesta i den här framställningen stämmer, och i flera avseenden tycker jag faktiskt att det här kapitlet är det mest läsvärda i hela boken. I och med att det handlar om en återgivning av händelser i amerikansk nutidshistoria och verifierbara uppgifter så finns det inte så mycket att förvränga. Filtrerar man bort den värsta negativa och stundtals sarkastiska jargongen så återstår en rätt bra överblick över ID-rörelsens organisation.
För att få perspektiv över E.s framställning i det här kapitlet beslöt jag mig för att sätta mig in i de dokument som han refererar till som någon sorts programförklaring för DI:s påstådda strategi för att ”ta över världen”, eller åtminstone det amerikanska samhället. De tre är (s 182, 184, 185):
1. The Wedge Document. DI:s informella programförklaring från 1999. 140
2. Roots: A grassroots network of Discovery Institute’s Center for Science and Culture. 141
3. The Changing of the Guard: Biblical Bluepronts for Political Action av George Grant, 1987.142
4. The Institutes of Biblical Law av Rousas John Rushdoony.143
Det här är resultatet av min efterforskning:
1. The Wedge
Wedge-dokumentet är kritiskt till den strängt naturalistiska utgångspunkten inom vetenskapen och dess destruktiva följdverkningar för det moderna tänkandet och samhället. Det ligger därför i sakens natur att E. är negativt inställd till det.
Men när man läser dokumentet med mer objektiva ögon framstår det klart att det inte är de negativa samhällskonsekvenserna av naturalismen som är dess huvudsakliga fokus, utan själva sanningsfrågan! När ID-rörelsens tidigare frontfigur Phillip E. Johnson skrev sin kända bok om strategin gav han den titeln The Wedge of Truth: Splitting the Foundations of Naturalism(2000). Dess centrala budskap är att evolutionsteorins brister som undanhålls den breda allmänheten måste, tillsammans med den positiva evidensen för en intelligent design, exponeras för allmänheten i första hand för sanningens skull. Just detta är också mitt eget fokus i det dokument du just nu läser. Sedan är det fullt sant att det här kommer att få positiva effekter för samhället på alla plan. Öppenhet och sanning är alltid framgångsfaktorer på både kortare och längre sikt.
Jag läste för många år sedan Phillip E. Johnsons The Wedge of Truth.144 Här ett citat från boken:
Man kan inte försvara vetenskapen genom att avfärda viktiga frågor, förlöjliga oliktänkande eller hindra människor från att ta del av bevisen för och emot de teorier som för närvarande råder. Den typ av vetenskap som förtjänar att försvaras är inte rädd för att möta kritik med sina egna metoder: välgrundade argument, precisa definitioner, repeterbara experiment och ett öppet sinne för alla frågor som inte kan avgöras genom opartiska vetenskapliga tester. Man kan försvara vissa teorier eller institutioner med andra metoder än de genuina vetenskapliga metoderna, men om man väljer den vägen kommer man så småningom att upptäcka att det man försvarar inte är vetenskap. (s 83, översättning DeepL)
E. skulle naturligtvis invända mot formuleringen ”precisa definitioner”, eftersom hans vederläggning av informationsargumentet går ut just på att biologisk funktionell information är svår att mäta. ID-teoretikerna å andra sida lägger ner stor möda på att lista ut teoretiskt och praktiskt hållbara sätt att mäta denna erkänt centrala biologiska storhet. Man kan fundera över vilket av de båda förhållningssätten som är mest konstruktivt ur vetenskapligt perspektiv.
The Wedge är därför en röst för öppenhet och sanning och därmed precis raka motsatsen till den antidemokratiska rörelse med dunkla avsikter som E. vill framställa den.
2. Roots
Roots, ”rötter”, är en resurssida för gräsrötter, det vill säga föräldrar, lärare och folk i allmänhet. Den erbjuder videolänkar, skriftliga sammanfattningar och allmänna tips om hur man kan sprida skapelsebudskapet i de sammanhang där man befinner sig. Dokumentet A Parent’s Guide to Intelligent Design har följande upplägg:
Del 1. Vad är Intelligent Design?
Del 2. Vad är evolution?
Del 3. Vilka är de vetenskapliga problemen med evolution enligt neodarwinismen?
Del 4. Hur kan föräldrar påverka evolutionsundervisningen i lokala skolor?
Del 5. Vad är teistisk evolution?
Del 6. Vad kan föräldrar göra för att stödja Intelligent Design?
Del 7. Grundläggande tips för föräldrar och studenter.
Del 8. Rekommenderade resurser.
Webbplatsen är överlag en bra resurs om man snabbt vill skaffa sig en överblick över vad ID-rörelsen står för. Personligen anser jag emellertid att föreningen Genesis är ett väl så bra alternativ för oss i Sverige. Jag har redan nämnt att jag/vi inte delar ID:s ställningstaganden till hundra procent, och jag återkommer strax till det.
Att E. anser Roots vara en aktör med den lömska avsikten att sprida villfarelser och underminera allmänhetens förtroende för vetenskapen får man naturligtvis se i ljuset av hans övertygelse att ID är lögn och ovetenskap. Själv kan jag inte se tendenser till någonting annat än en strävan efter öppenhet och en saklig och allsidig undervisning i materialet. Jag säger som Charles Darwin själv gjorde i sin klassiska bok Om arternas uppkomst:
ett rättvist resultat [om ursprungsfrågan] kan endast uppnås genom att fullständigt redogöra för och väga fakta och argument från båda sidor i varje fråga.145
Det där borde faktiskt E. ta lärdom av.
3. The Changing of the Guard
Jag har av tidsskäl bara läst delar av dokumentet The Changing of the Guard som E. citerar med bland annat orden
Det är herraväldet vi vill ha (s 185)
Men jag bad ChatGPT om en saklig sammanfattning av dokumentet. Det visade sig stämma väl överens med det intryck jag själv fick:
Det argumenterar för att kristna bör arbeta för ett samhälle präglat av bibliska värderingar genom politiskt och kulturellt engagemang, utan att formulera ett detaljerat religiöst rättssystem.
Författaren till Changing of the Guard är kritisk mot modern värdeneutral demokrati, men förespråkar inte avskaffande av demokratiska processer. Snarare principer som vore naturliga i ett samhälle där en majoritet vill leva efter bibliska principer i enlighet med hur Nya testamentets författare presenterar det kristna budskapet.
Viktigast i sammanhanget är förstås den av E. påstådda kopplingen till ID-rörelsen. Jag ställde följdfrågan till AI-assistenten om det framkommer någon sådan i boken. Jag fick följande svar:
Sekundära sökningar av bokens innehåll och sammanställningar av verk som ingår i samma serie visar inte heller någon koppling till diskussioner om Intelligent Design eller relaterade vetenskapliga begrepp. De visar snarare att The Changing of the Guard fokuserar på kristen politisk strategi och dominionteologi (ansvaret att etablera kristen lag och ordning i civil samhällsordning), men inte på ID eller evolution/ursprungsdebatter. (min fetmarkering)
4. The Institutes of Biblical Law
E. har däremot rätt i att dokumentet The Institutes of Biblical Law av Rousas John Rushdoony diskuterar en extrem form av rekonstruktionism som utgår från det Gamla testamentets lagar och huruvida de bör tillämpas i juridiska sammanhang i händelse av att ett land (läs: USA) skulle få en kristen politisk majoritet. Det är en fråga som leder till ett spänningsfält mellan teokrati (Gud bestämmer) och demokrati (folkmajoriteten bestämmer) som säkert är jätteintressant, men som jag inte skulle ge mig in i därför att den kräver en helt annan nivå av både vilja och kompetens än jag har.
Men det som emellertid är av intresse även här är givetvis den koppling till ID-rörelsen (eller kreationism) som går att utläsa i det här drygt tusensidiga dokumentet. Det är ju i det sammanhanget som E. tar upp det. Återigen rådfrågade jag ChatGPT och fick ett svar med följande sammanfattning (observera att det fetstilta återigen är ChatGPT:s eget val):
Intelligent design-rörelsen nämns inte i The Institutes of Biblical Law, Vol. 1 och det finns inget i boken som diskuterar denna rörelse eller begreppet ”intelligent design” i den moderna betydelsen. Det finns alltså ingen relevant kontext i denna bok där begreppet förekommer.
I texten omedelbart före sammanfattningen sägs vidare:
Innehållsförteckningen för boken listar kapitel om lagens natur, budorden, brott och straff, familj, egendom, domstolar, etc., men ingen del rör vetenskapsteori, naturvetenskap, eller begrepp som intelligent design.
Jag är (som alltid) helt öppen för att låta mig motbevisas av någon som hävdar motsatsen, men bara av faktisk evidens, inte av diverse tyckanden.
Discovery Institute och rekonstruktioniströrelsen
Om det nu inte går att koppla ID till den rekonstruktionistiska rörelsen i USA, så kanske det går att koppla Dicovery Institute som organisation till den? E. namnger ”en tidigare ledande rekonstruktionist, multimiljonären Howard Ahmanson Jr” som finns med i DI:s ledning. Eftersom jag aldrig hade hört namnet tidigare bestämde jag mig för att återigen anlita ChatGPT. Jag frågade helt enkelt:
Stämmer det att en viss Howard Ahmanson Jr är en tidigare ledande rekonstruktionist?
ChatGPT svarade:
Formuleringen är problematisk. Ahmanson var inte en teoretisk ledare för rörelsen. “Ledande rekonstruktionist” antyder normalt att man är en central ideolog eller organisatorisk huvudfigur. Inom kristen rekonstruktionism brukar man i stället räkna t.ex.: Rousas J. Rushdoony (grundare), Gary North och Greg Bahnsen. Ahmanson räknas inte till rörelsens teologiska eller organisatoriska ledarskap. Han var främst finansiär och sympatisör. Det som är väl belagt är att han:• finansierade projekt kopplade till Rushdoonys krets (Chalcedon Foundation)
• uttryckte sympati för delar av idévärlden under 1970–80-talen
• senare tog viss distans från delar av läran
Detta gör honom mer till donator och supporter än ledare. Han har själv tonat ned kopplingen senare. I senare intervjuer har han sagt att han inte delar alla rekonstruktionistiska ståndpunkter, vilket gör etiketten ännu mer osäker.
Visst, man kan inte alltid lita på AI-genererade svar, men min erfarenhet är att de brukar vara ganska bra på just den här sortens frågor. Återigen – bevisa gärna motsatsen.
Tyvärr blir det min slutsats att E. handskas ovarsamt med fakta. Genom att utan att ha täckning för det han påstår sammankoppla ID-rörelsen med politisk dominionism/rekonstruktionism bidrar han till att skapa en lögn och en konspirationsteori genom guilt-by-association. Det här är allvarligt! Man kan begära mer än så av en företrädare för vetenskapen som E.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 27. Tre centrala misstag
Här tar E. upp misstag som han anser att ID-rörelsen gör.
1. Att etik och moral måste ha sin grund i religionen, och speciellt kristendomen.
Det här har jag mig veterligen aldrig noterat, trots att jag följt rörelsen sedan den startade. Vad jag däremot läst återkommande gånger – och som jag också är övertygad om stämmer – är att när ateister och agnostiker refererar till sin egen etik och moral, så går det inte att hitta någon förankring för någon viss sådan i deras egen världsbild. Man kan som ateist absolut tycka att det är fel att stjäla, mörda och leva rövare och verkligen leva därefter. Men det går knappast att hitta de begreppen som några dygder i evolutionsteorin som sådan, möjligen i en och annan mer eller mindre godtycklig tolkning av den. I en naturalistisk/evolutionär världsbild existerar etik och moral av den enda anledningen att individer med sådan fått större reproduktionsframgång, inte för att de skulle ha någon principiell giltighet. Jag rekommenderar magasinet Genesis temanummer om Världsbilder.146
2. Att tro att man kan bedriva naturvetenskap med övernaturliga förklaringsmodeller.
Det här har jag redan avhandlat i kapitel 12.
3. Att tro att vetenskapen är ateistisk, det vill säga inte kunna skilja mellan metodologisk och filosofisk naturalism.
Också detta är en vinklad formulering. Ingen ID-teoretiker eller kreationist skulle formulera sig så. Jag drar mig till minnes följande ord av William Dembski:
Vi behöver inse att den metodologiska naturalismen är funktionellt likvärdig med en fullfjädrad metafysisk naturalism. Den metafysiska naturalismen hävdar att den materiella världen är allt som finns. Den metodologiska materialismen vädjar till oss om att för vetenskapens skull låtsas som att den materiella världen är allt som existerar. Men i det ögonblick som naturvetenskapen anammas som den enda universellt giltiga formen av kunskap i en kultur, följer omedelbart att metodologisk och metafysisk naturalism i alla avseenden blir omöjliga att särskilja. De är funktionellt likvärdiga.147
Min personliga hållning är att kristna naturvetare bör sluta omfamna och försvara sträng metodologisk naturalism som kan låsa fast forskningen på blindspår. I stället bör man förespråka pragmatisk naturalism,148 som tillåter avsteg från den övergripande principen om metodologisk naturalism när situationen så kräver.
Det finns, som vi tidigare har berört (kap 10) olika strategier som evolutionister brukar alternera med att använda i sin polemik mot oss anhängare av skapelseperspektivet. I den här boken framför allt två.
Den ena är en offensiv, militant variant, som handlar om att ”debunka” våra argument och visa hur urbota korkade vi är som missförstår vetenskap i allmänhet och evolutionsteorin i synnerhet. Som läsare bör du vid det här laget ha insett att det är E.s huvudstrategi.
Den andra strategin är av en defensiv karaktär. Den handlar om att övertyga troende personer om att intelligent design och kreationism är dåliga alternativ för kristna (tangeras direkt eller indirekt på s 8, 78, 199) och att det finns ett bättre alternativ i teistisk evolution (s 72). Framlidne Stephen J. Gould kallade denna strategi NOMA (Non-Overlapping Magisteria). Det skulle alltså vara att acceptera evolutionsteorin med hull och hår medan man går med på att hålla tron på Gud på det privata planet utan några anspråk på att Han skulle ha någonting med den fysiska världen att göra.
Religiösa människor med överdriven respekt för ”vetenskapen”, och i synnerhet naturvetenskapen, löper stor risk att hamna där. De personer (varav ett flertal präster i Svenska kyrkan) som medverkar i samma dialoggrupp som E. befinner sig sannolikt där.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 28. Intelligent design som gudsbevis
I detta korta kapitel har E. också en poäng. Han konstaterar (s 200) att ID-rörelsen vill framstå som en religiöst neutral part som inte bygger designperspektivet på någon helig skrift som Bibeln. Precis som han påpekar är rörelsen negativt inställd till teistisk evolution, som exempelvis förespråkas av Francis Collins och den teistisk-evolutionistiska propagandaorganisationen BioLogos som han grundat. Det framgår också av antologin Theistic Evolution: A Scientific, Philosophical and Theological Critique.
Det här skapar en viss tvetydighet inom ID-rörelsen som jag personligen inte tror är så sund. Det kan vara en grogrund för kommande splittring inom rörelsen, vilket vore olyckligt. ID-rörelsen är idag dessutom öppen för idén om gemensamt släktskap,149 en syn som vi i föreningen Genesis absolut inte delar.
Ljuset i det här dunklet är att det finns en tendens att personer inom ID de senaste åren rört sig i riktning mot ungjordskreationism. Jag ser det med tillfredsställelse, för jag anser att det är det mest stabila viloläget där man minimerar konflikter både på teologins område och i relation till den naturvetenskapliga evidensen. Men naturligtvis inte med företrädare för den evolutionära berättelsen som dominerar den sekulära akademiska världen, däribland bokens författare.
↩ Tillbaka till innehållsförteckningen
![]()
Kapitel 29. Intelligent design som politisk agenda
Vi har nu nått fram till bokens sista kapitel. Det kan sammanfattas i citatet:
[Vetenskapen har inte] längre den respekt och den höga status den tidigare haft. Discovery Institute rider på den här vågen av antivetenskaplig populism och förstärker den ytterligare. (s 202)
Problemet som E. beskriver är reellt. E. försöker projicera det på ID, men som jag påpekade i min kommentar till det intuitiva argumentet (s 83 ovan) så är det frågan om det inte snarare är den evolutionära dogmatismen och den akademiska tystnadskulturen överlag som är roten till det onda, i kombination med samhällets postmoderna strömningar.
Återstår gör bara epilogen.
↩ Tillbaka till innehållsförteckningen
![]()
E. skriver:
De vetenskapliga idealen om rationellt, empiriskt och logiskt tänkande fritt från mytiska förklaringar är i stor utsträckning ett arv från den antika grekiska filosofin. De krafter som nu lanserar ID vill att vi ska återinföra högre makter som förklaring i det vetenskapliga arbetet. Det är ett försök att vrida klockan 2 500 år tillbaka. (s 204)
Han lovprisar ”rationellt, empiriskt och logiskt tänkande fritt från mytiska förklaringar”. (s 204) Jag har i den här bokanalysen visat att E. själv brister på samtliga dessa punkter. Inte bara i något enstaka fall, utan vid så många tillfällen att jag saknar ork att räkna upp dem alla. Det vapen han drar mot ID och kreationism kan alltså vändas mot honom själv.
Och ja, modern vetenskap har visst ett arv från antiken. Men E. glömmer att det inte var där och då som vetenskapen utvecklades. Läs gärna om anledningen i Appendix 4.
↩ Tillbaka till innehållsförteckningen
![]()
Den amerikanske evolutionsbiologen Richard C. Lewontin sade en gång: “…vi får inte tillåta en gudomlig fot i dörrspringan”. Det har också varit också E.s ambition med den här boken.
Tyvärr har han i sin iver att gå i Lewontins fotspår försatt sig i en situation där han berättar halvsanningar, utelämnar viktiga fakta och ibland (medvetet eller omedvetet må vara osagt) rena faktafel och falska beskyllningar, samtidigt som han både på och mellan raderna för fram sin egen reduktionistiska verklighetsuppfattning. Återigen: Det är han i sin fulla rätt att göra. Det blir bara lite ironiskt när han å ena sidan beskyller ID-rörelsen för ovetenskaplighet och smygreligiositet därför att de för in designbegreppet i vetenskapen och på samma gång själv öppet förordar en reduktionism med tydliga naturmagiska tendenser som om den vore vetenskapligt förankrad.
E.s avsikt har varit att visa hur förträfflig evolutionsteorin och dess förutsättning metodologisk naturalism är, hur usla argument designrörelsen och kreationister har, och vilka allvarliga konsekvenser dessa kan ha för samhället. Nu är den föreställningen inte längre oemotsagd, och jag vill uppmana dig som läser det här dokumentet att sprida det på alla sätt du kan!
Den naturalistiska världsbilden med evolutionen som sin ”skapelseberättelse” brister som vi sett i förklaringskraft. Det gäller inte bara i fråga om naturens strukturer och funktioner och dess underliggande informationssystem, utan även allt det andra som hamnar utanför när locket på naturalismlådan är stängt, som till exempel det mänskliga medvetandet, nära-döden-upplevelser, skönhet, etik och moral.
Minns du de tre formuleringarna i bokpresentationen på Bokus?
• ID-rörelsen är en del i ett kulturkrig mellan liberal demokrati och religiös fundamentalism.
• Boken synar noga argumenten för intelligent design och visar varför de inte håller.
• En bok om varför det spelar roll vad vi bygger vår världsbild på.
Jag nu visat att den första punkten är helt grundlös (kap 26).
Jag har granskat E.s kritik med avseende på samtliga tretton argument (kap 12-24) och visat att den bygger på missförstånd och faktafel. Inte heller den andra punkten stämmer alltså. I förordet till boken (s 8) uttrycker författaren ett särskilt tack till professor Dan Larhammar för hans ”sakkunniga läsning av texten”. Det innebär att jag tvingas ge samma underkända betyg även till honom. E. riktar också tack till undertecknad, Ola Hössjer och Mats Selander för de diskussioner vi haft med honom på nätet. Det har ”varit till stor hjälp” skriver han, eftersom han ”haft svårt att förstå hur ID-förespråkare resonerar”. Det är uppenbart att vi inte nått fram med vårt budskap. Kommer den här motboken kommer att lyckas bättre?
Och även den tredje punkten är i grunden fel, eftersom det förhåller sig precis tvärt om. Vi bygger inte vår världsbild på en samling fakta och logiska resonemang. I stället utgår vi från en världsbild när vi tolkar fakta och för våra resonemang. Det är E.s personliga val av världsbildsglasögon som gör honom fullt och fast övertygad om evolutionens faktum, dess gradualistiska mekanismer och ID-argumentens tillkortakommanden. Det är tack vare dessa glasögonfilter som han inte kan se att mutationer är djupt destruktiva och förstår att selektion utarmar. Det är tack vare dem han inte ser vilken överväldigande evidens för en Designer som de tretton argumenten representerar.
Har då inte jag själv några glasögon? Visst har jag det. Min världsbild är den bibliska, där Guds existens inte ens behöver motiveras. Betraktad genom de glasögonen är den uppenbar på alla nivåer i skapelsen.
Det som skiljer mitt och E.s perspektiv är därför inte i första hand kunskap och erfarenheter, det har vi gott om båda två, och till och med delvis inom samma områden. Det som skiljer oss åt är våra världsbilder. Vilken världsbild som är den vanligaste i akademiska och många andra sammanhang vet vi redan, det är den naturalistiska. Men det är faktiskt inte det som avgör saken. Den centrala frågan är vilken av våra världsbilder som är den mest trovärdiga. Det ligger nära till hands att tro att den mest trovärdiga världsbilden också är den mest sanna.
Men den frågan ligger utanför ramen för den här diskussionen. Kanske återkommer jag till den i ett annat sammanhang. Jag rekommenderar läsaren att börja nysta i världsbildsfrågan. Varför inte börja den resan med att läsa magasinet Genesis temanummer om Världsbilder (not 146).
Låt mig avsluta med tre citat. Det första av den ateistiske filosofen Thomas Nagel,150 och de två sista från en tusentals år gammal bok:
Jag talar av erfarenhet och är starkt utsatt för den här rädslan själv: Jag vill att ateismen skall vara sann, och jag blir obehaglig till mods av det faktum att några av de mest intelligenta och välinformerade personer jag känner är religiösa troende. Det är inte bara det att jag inte tror på Gud och naturligtvis hoppas att jag har rätt i min tro. Det är det att jag hoppas att det inte finns någon Gud! Jag vill inte att det ska finnas någon Gud; jag vill inte att universum skall vara sådant. Min gissning är att detta kosmiska auktoritetsproblem inte är något sällsynt tillstånd och att det är orsaken till mycket av vår tids scientism och reduktionism. En av de tendenser det ger stöd åt är den löjliga överanvändningen av evolutionsbiologi för att förklara allting som rör det mänskliga livet, inklusive det mänskliga medvetandet.
***
Ska inte örat pröva orden, liksom munnen prövar matens smak? (Job 12:11)
Den som först för sin talan har rätt, tills motparten kommer och prövar saken. (Ord 18:17)
↩ Tillbaka till innehållsförteckningen
![]()
Nära-neutrala mutationer och det naturliga urvalet
Problemet med nära-neutrala mutationer är att deras effekt på organismen är så obetydlig att det naturliga urvalet inte “upptäcker” dem, eftersom de bildligt talat dränks i “bruset” av en uppsjö andra faktorer.
Låt oss rent hypotetiskt tänka oss två personer där den ene (A) fått en svagt negativ mutation som sänker hans synskärpa en liten aning, så litet att det nätt och jämnt märks av vid en synundersökning. Den andra (B) får en av de där extremt sällsynta fördelaktiga mutationerna som ger henne en aning bättre synskärpa, som även den nätt och jämnt är upptäckbar hos ögondoktorn.
Kommer nu selektionen att gynna B framför A? Om vi tänker bort alla andra faktorer som kan påverka en människas levnad så finns det en ytterst liten teoretisk möjlighet att det skulle påverka fortplantningsframgången lite till det sämre för A och aningen bättre för B.151
Men så har vi det här med verkligheten. Person A kan av en ren tillfällighet ha ärvt gener för lite extra vackra ögon, eller någon procent mindre risk för hjärt- och kärlsjukdomar av sina föräldrar. Eller han kan vara född i ett land där medellivslängden är ett halvår längre. Så där kan man hålla på. I verkligheten finns det oräkneliga faktorer som konkurrerar med den där ögongenen och som gör att selektionen inte kommer åt att eliminera A.
På motsvarande sätt kan person B med aningen bättre synskärpa ha ärvt en gen för typ 2-diabetes eller ha en ovana att äta för mycket chips eller helt enkelt ha otur när han skulle gå över trottoaren…
I den nämnda boken Genetic Entropy illustrerar John Sanford detta med H C Andersens saga om prinsessan på ärten. En liten flicka som ligger på en fyra meter tjock madrass ovanpå en ärta. Det är bara i sagans värld som hon kan uppleva obehaget av ärtan. I verkligheten är det lika omöjligt som det är för det naturliga urvalet att “känna “av nästan-neutrala mutationer, vare sig de är lite bra eller lite dåliga.
↩ Tillbaka till innehållsförteckningen
![]()
Kiralitet
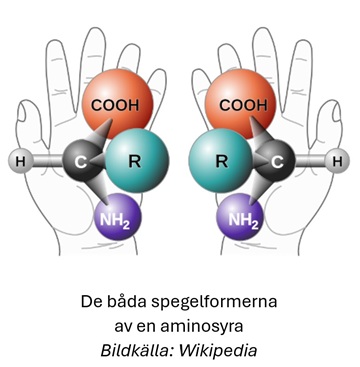
Även om det egentligen inte finns någon grundläggande skillnad mellan livlös och levande materia så finns det en speciell egenskap som faktiskt är unik för de ämnen som finns i de levande varelserna: De är nästan alltid utpräglat asymmetriska, eller som det också heter – kirala. Namnet kommer från grekiskans kheir som betyder ”hand”. Om man skulle be eleverna i en skolklass göra molekylmodeller av en aminosyra skulle ungefär hälften av eleverna montera ihop atomerna på ett sätt, och den andra hälften på ett annat. Båda är rätt. Skillnaden är att de båda är varandras spegelbilder.
Fenomenet kallas spegelisomeri och studeras inom ett särskilt område av den organiska kemin som kallas stereokemi. Det har viktiga tillämpningar inom bland annat läkemedelsindustrin152. Situationen uppstår på grund av att kolatomen (med beteckningen C i figuren) binder till fyra olika atomgrupper.153 De två spegelbildsformer av aminosyrorna – eller vilket slag av molekyl det gäller – brukar kallas L- respektive D-formen154.
Faktum är att det var den berömde franske vetenskapsmannen Louis Pasteur som var pionjären inom detta område. Han formulerade det rentav som en naturlag att levande varelser, till skillnad från mineralvärlden, består av asymmetriska molekyler. Det visade sig inte stämma. De enskilda (ofta mikroskopiska) kristaller som bygger upp olika mineral, som till exempel kvarts (SiO2), är antingen av L- typ eller D-typ, men fördelningen dem emellan är 50:50.
Proteiner är den ämnesgrupp som ger allt levande dess form och funktion. Formen genom så kallade strukturproteiner och funktionen oftast genom enzymer som påverkar och reglerar cellens alla kemiska reaktioner – dess ämnesomsättning. Kemiskt sett är proteiner kedjor av aminosyror. Antalet aminosyror (eller formellt ”aminosyrarester”) varierar från några tiotal upp till tiotusen. Men alltid bara L-formen av de enskilda aminosyrorna. Man säger att proteinerna är homokirala med avseende på sina aminosyror. Detta är anmärkningsvärt, eftersom tillverkning av aminosyror alltid leder till blandningar med 50% av varje slag. Det är till och med så, att om man löser upp ren155 L- eller D-form av en aminosyra i lite vatten i ett provrör, så kommer det efter en stund att finnas lika mycket av båda formerna i röret – kemister kallar det då för en racemisk blandning eller ett racemat. Att det trots detta bara finns den ena sorten i alla156 proteiner beror på att det finns speciella enzymer i cellerna som ser till att det bara blir den ena av dem som monteras. Med andra ord ser homokirala proteinmolekyler till att det bara bildas homokirala proteinmolekyler i de levande varelserna. Det påminner inte så lite som den klassiska gåtan om vilket som kom först – hönan eller ägget…
Samma sak gäller för de sockerarter som ingår i nukleinsyrorna DNA och RNA som hanterar lagring och överföring av information i cellen, men med skillnaden att det där bara förekommer D-formen av sockermolekylerna. I praktiken är problematiken ännu större med dessa sockermolekyler. Det finns till exempel åtta olika men snarlika sockermolekyler som ribos, men bara en av dem förekommer i RNA. Hur kunde det bli så, kan man undra. Hur det än gick till är forskarna eniga om att det måste ha skett i samband med att den första levande cellen uppstod, eftersom det är en gemensam egenskap för alla jordens livsformer.
Homokiraliteten är så viktig att det räcker med en enda aminosyra av fel variant för att en proteinmolekyl helt ska förlora sin 3D-form och därmed sin funktion. Och detsamma gäller i DNA-molekylen – en stackars L-sockermolekyl bland hundratusentals D-molekyler i DNA-spiralen och molekylen skulle få en konstig böj som skulle göra att den slutade fungera som informationsbärare.
Att ur en blandning av lika mängder L- och D-molekyler slumpmässigt bilda en kedja av 300 molekyler av enbart den ena varianten (vilket motsvarar längden på ett medelmåttigt protein och en kort nukleinsyra) skulle vara som att singla slant och få samma sida av myntet uppåtvänt 300 gånger i rad. Pröva själv med att lyckas 10 gånger så inser du problemet för teorin om hur proteiner, DNA och RNA uppstod spontant. Om man skulle använda matematisk statistik för att räkna på detta så inser man snart att slumpen ensam omöjligen kan vara orsaken. Sannolikheten att ur en ocean av en racemisk blandning av aminosyror slumpmässigt få fram en sekvens med 300 L- (eller D-)aminosyror kan liknas vid att ur en ocean av stora och små bokstäver (versaler och gemener) med förbundna ögon råka plocka upp 300 versaler i rad. Sannolikheten för det är 50 procent (½) varje gång, och sammantaget (½)300. Utför man den beräkningen på en miniräknare får man som resultat ett tal som är en chans på 2 x 1090. Talet skrivs ut som en tvåa följd av 90 nollor, och är 20 miljarder gånger fler än det sägs finnas elementarpartiklar i universum.
Av detta kan man dra den enkla slutsatsen att det måste finnas andra orsaker än slumpen till livets asymmetri.157 Forskare har under åren föreslagit ett antal olika hypoteser om hur det skulle kunna ha gått till. En av dem handlar om att en speciell form av polariserat ultraviolett ljus skulle kunna ha orsakat att det bildades mer av den ena eller andra spegelbildsformen. Kruxet är att den sortens ljus hittills bara lyckats framställas på teknisk väg och användas på laboratoriet. Andra förslag har varit lermineral med viss förmåga att gynna den ena spegelformen. Även starka magnetfält och en viss typ av radioaktivt sönderfall har föreslagits. Inget av dessa alternativ har dock visat sig kunna leda till så pass stark asymmetri att det skulle bli statistiskt rimligt att förvänta sig några längre kedjor av bara den ena sorten. Kom ihåg att det räcker med en enda felaktig byggsten någonstans i kedjorna för att förstöra alltsammans, åtminstone i dagens värld, men någon annan värld än denna är det ju svårt att uttala sig om. Homokiraliteten hos livets molekylkedjor kvarstår därför som en av ursprungskemins allra största utmaningar.
Men tänk dig att du återigen står framför den där oceanen av stora och små bokstäver. Tänk dig vidare att du den här gången tar av dig ögonbindeln och använder dina sinnen till att aktivt välja ut 300 bokstäver som alla är versaler. Och kanske dessutom rentav skapar en liten trevlig berättelse av dem. Det skulle inte ta många minuter. Tänk vilken oerhörd förklaringskraft som fenomenet intelligens och målmedvetenhet har i relation till slumpen. Det är jämförbart med Dawkins vessla som du läste om i kapitel 16. En intelligent design, en gudomlig skapelseakt, är därför inte bara en högst relevant – utan i praktiken den enda realistiska – förklaringen till homokiraliteten hos livets molekyler och den genetiska information den bär.
↩ Tillbaka till innehållsförteckningen
![]()
Naturens skönhet
Här följer en redogörelse för hur biologer har försökt förklara naturens skönhet ur ett evolutionsperspektiv. Fokuset är på skönheten hos levande varelser. Jag presenterar förklaringsmodellerna i en någorlunda kronologisk ordning, även om de delvis existerat samtidigt. Texten är tidigare publicerad i Genesis magasin med temat Skapelsen och skönheten.158
1. Charles Darwin – sexuell selektion
Charles Darwin själv var väl medveten om att naturens skönhet pockade på en förklaring, och tyckte att påfågelns fjäderdräkt gjorde honom illa till mods. Han brottades med frågan och elva år efter publiceringen av sitt klassiska verk ”Om arternas uppkomst” lade han fram sin teori om sexuell selektion. Syftet var att förklara djurvärldens så kallade ”sexuella ornament” – det vill säga vackra utstyrslar hos framför allt hannar – inklusive påfågeltuppar.
Darwins teori gick ut på att påfågelhönorna helt enkelt valde ut den av de uppvaktande tupparna som hade de vackraste stjärttäckarna. När denna urvalsprocedur upprepades generation efter generation ledde det så småningom till tupparnas nuvarande utseende.
Men att vara mer iögonfallande och otympligare i sina rörelser på grund av extremt långa stjärtfjädrar innebar förstås samtidigt en successivt ökande risk för påfågeltupparna att bli tagna av ett rovdjur. Den nackdelen, ansåg Darwin, kompenserades mer än väl av att de vackraste exemplaren hade större framgång vid sina frierier. Darwins samtida kritiker påpekade att detta förutsatte att hönorna skulle ha ett liknande skönhetsideal som vi människor, vilket de ifrågasatte.
2. Skönhet som fitnessmarkör
Enligt denna syn avspeglar skönheten djurens fitness.159 Sexuella ornament i form av en ovanligt vacker fjäderdräkt skulle alltså attrahera honorna därför att det signalerade saker som motståndskraft mot sjukdomar och parasiter eller en god näringsstatus. Genom att påfågelhönorna valde hannarna med de vackraste fjädrarna valde de därmed automatiskt även de friskaste och starkaste exemplaren. Detta har länge varit den dominerande synen bland evolutionsbiologer. Hypotesen förutsätter dels en fysiologisk koppling mellan skönhet och bra fitness, och dels att hönorna intuitivt inser detta.
En variant av denna hypotes har en mer indirekt koppling mellan graden av ornament och fitness. Enligt detta synsätt proklamerar den hanne som står där med en extrem ”utstyrsel” med sin blotta existens budskapet att han mår väldigt bra – något i stil med: ”Trots alla de handikapp som min utstyrsel inneburit för mig av ökade risker och högre energiförbrukning har jag ändå lyckats komma undan mina förföljare och konkurrenter och står här nu inför dig i all min prakt – välj mig!” Även denna variant förutsätter naturligtvis att hönorna fattar galoppen.
3. Miljöfaktorer och tillfälligheter startar utvecklingen av egenskaper
Enligt det här modernare synsättet tänker man sig att en ren tillfällighet eller någon detalj i miljön kan trigga igång en förkärlek hos en hona för någon mer eller mindre godtycklig egenskap hos en hanne, som till exempel en viss färgnyans, längd på antenner, fjädrar eller någonting annat. Sedan tänker man sig att denna svaga tendens till förkärlek för någonting startar en selektionskedja som så småningom leder till utpräglade uttryck av dessa egenskaper, som en intensiv färg eller extremt långa antenner eller fjädrar. Enligt det här synsättet är det inte bara den fysiska miljön som formar organismernas skönhet genom sitt ”selektionstryck”, utan i minst lika hög grad organismernas egna subjektiva upplevelser av den – deras ”sensoriska bias”. Även denna hypotes förutsätter samma estetiska urskiljningsförmåga hos djuren som de tidigare varianterna.
4. Idag - allt på samma gång
En allt vanligare hållning bland dagens evolutionsbiologer förefaller vara någon sorts blandning av alla de här synsätten – inte något ”antingen eller” utan snarare ett ”både och”. E. nämner neutral evolution och sexuell selektion. Vid sexuell selektion är det djurens – och framför allt honornas estetiska sinne som är nyckeln.
Finns det vetenskaplig evidens för att ett sådant existerar? Svar: Nej.160
5. En fråga om evidens
Anledningen till den här ganska spretiga evolutionära synen på naturens skönhet är nämligen just bristen på evidens. Det har utförts en mängd experiment i syfte att styrka de nämnda hypoteserna, men resultaten har varit nedslående.161 Observationer av hur exempelvis påfågelhönor väljer sina partners ger inget stöd för idén att de väljer tupparna med flest och snyggast stjärtfjädrar.162
De evolutionistiska förklaringarna förutsätter saker som inte alls är självklara. Till exempel följande:
• Varför skulle honor av olika djurarter ha samma estetiska sinne som vi?163
Även om det kunde visas att påfågelhönorna verkligen hade föredragit tuppar med vackrare stjärtpennor, skulle det inte förklara hur denna ändamålsenliga ”attityd” hos hönorna uppkommit. Darwins förklaring av hannarnas skönhet i djurvärlden bygger på att organismer (framför allt honor) har sina egna uppfattningar om estetik och skönhet som påfallande ofta råkar sammanfalla med våra egna värderingar.
Idén om skönhet som fitnessmarkör förutsätter att honorna av evolutionen utrustats med en intuitiv insikt om att hannarnas skönhet visar på deras fitness i form av styrka, sundhet, fertilitet och intelligens, och att det är därför de väljer de vackraste hannarna. Det förutsätter en algoritm som kopplar skönhet till fitness i en positiv korrelation (d v s ”ju mer välanpassad desto vackrare” och inte ”ju mer välanpassad desto fulare”).
Det är förstås inte alls självklart att en i vårt tycke vacker färg eller vackert mönster skulle uppfattas på samma sätt av en djurhona. En doft som för oss är vidrig kan ju till exempel ofta uppfattas som mycket tilltalande av vår hund. Det kunde alltså lika gärna ha varit precis tvärtom, att honorna attraherades av den hanne som hade de i vårt tycke tristaste fjädrarna. Vore det slumpmässiga processer som styrde kunde man förvänta sig att det i ungefär hälften av fallen faktiskt vore så, och då hade den totala nettotendensen till estetisk skönhet i världen – så som vi uppfattar den – ha varit lika med noll. Antag att det verkligen skulle finnas en ”skönhetsalgoritm” inbyggd i honornas sinne (det vet vi inte säkert) – hur hamnade den i så fall där?
• Hur uppkom egenskaperna från början?
När påfågelhönan väl fått en förkärlek för en viss egenskap i tupparnas stjärttäckare, t ex fjädrarnas längd, antalet ögon, deras symmetriska placering eller färgsättning, skulle sexuell selektion teoretiskt sett kunna driva en förändring mot mer utpräglade egenskaper. Men det förklarar inte hur de olika länkarna i den komplexa tillverkningskedja som leder fram till de geometriska mönstren och färgsättningarna på fjädrarna uppkommit – och inte heller de genetiska programvarorna som kodar för dem. Detta är en helt annan fråga. Återigen – darwinismen kan förklara ”survival of the fittest” men inte ”arrival of the fittest”.
• Varför sådant överdåd av skönhet i naturen?
Vid första anblicken kan man tycka att det vore rimligt med en koppling mellan å ena sidan skönhet och å andra sidan fitness - man är ju vackrare när man är frisk än när man har influensa. Men kanske är kopplingen inte så självklar trots allt. Det borde nämligen finnas en någorlunda balans mellan mängden skönhet i naturen och mängden tillgängliga ”fitnessfaktorer” som de skulle svara emot. Rent intuitivt inser de flesta av oss att naturens till synes godtyckliga skönhetsaspekter är fullständigt överväldigande. Det räcker att skriva in orden ”beauty” tillsammans med ”insects”, ”birds, ”fish” eller ”flowers” och sätta sökmotorn i bild-läge så behöver vi inte längre betvivla den saken.
• Varför så ojämlikt?
Lövsalsfåglarnas parningsritual består av en hel arsenal av estetiska uttryck koncentrerade i en ceremoni som talgoxar och gråsparvar tycks klara sig alldeles utmärkt förutan. Ändå existerar de senare i högsta välmåga. Varför? Vilken är den evolutionära ”förklaringen”? Svar: Det bara råkade bli så. Lite liknande resonemang som att vissa avkomlingar av den första bakterien på jorden utvecklades till människor medan andra förblev bakterier ända in i våra dagar.
• Inte bara komplexa djur
Att en påfågelhona eller lövsalsfågelhona med sina avancerade nervsystem kan ha ett estetiskt sinne är kanske inte helt osannolikt, men insekter? Med tanke på alla vackra småkryp har tydligen även de haft liknande uppfattningar som oss om vad som är tilltalande när det gäller formgivning och färgsättning, och tycks även ha kunnat kvantifiera skönheten genom att ha kunnat välja den vackraste av sina partners under ”årmiljonerna”.
För att inte tala om blommors skönhet. Hur förklarar evolutionsteorin hela den botaniska världens mångfald av skönhet i former, färger och dofter? Svar: Som en ”samevolution” mellan pollinerande insekter och växterna. Men betyder det att bin och blomflugor har samma estetiska sinne som vi, de också? Vore det inte ganska märkligt?
Evolutionistiska bortförklaringar och fantasier
Hur hanterar evolutionsbiologer observationer som att påfågelhönorna inte verkar ett dugg intresserade av tupparnas fjäderdräkt? Svar: Genom att konstatera att det förr i tiden måste ha förhållit sig annorlunda. Idag när fjäderdräkten nått sin perfektion har evolutionen avstannat och hönorna förlorat sin ”selektionskompetens”. Verkliga observationer väger uppenbarligen inte särskilt tungt när de ställs mot tron på ”evolutionens faktum”. Ännu en gång: evolutionsteorin förklarar både allting och ingenting, både X och -X.
Filosofen Denis Dutton har argumenterat för att människor generellt har en medfödd uppskattning av en speciell sorts landskap – ett gräsbevuxet fält med dungar av träd, vatten och vilda djur – därför att det påminner om de savanner där människan evolverade under Pleistocentiden. I ett TED-talk164 förklarar han att evolutionen är anledningen till att konstnärer genom historien avbildat just den typen av landskap. Anledningen till att de avbildar träd som förgrenar sig nära marken är att våra förfäder förlitade sig på dessa bekvämligt låga grenar för att klättra upp i och klara sig från predatorer. Berättarkonst inspirerad av tron på samma ”evolutionens faktum”, men inte mer än så. Det går givetvis lika bra för en Bibeltroende att hävda att det där är ett nedärvt ”minne” från Edens lustgård. Vem vet?
En alternativ förklaring
De centrala nervsystemen hos olika grupper av djur skiljer sig avsevärt från människans, i synnerhet om man jämför små fågel- eller bihjärnor med våra egna. Därmed bör deras förmåga att uppfatta estetik och skönhet rimligen vara väldigt annorlunda än vår. Men den evolutionära förklaringen till skönhet bygger på att denna verkligen kan uppfattas, och inte nog med det, dessutom rangordnas av organismerna (framför allt honorna) så att de vackraste och mest välanpassade individerna väljs ut i varje generation. Det finns ingen som helst vetenskaplig evidens för att detta skulle vara möjligt, i synnerhet inte när det gäller mer enkelt byggda organismer.
Men ändå är hela djurvärlden, liksom även växtriket och svampriket, präglat av en mängd skönhetsdimensioner. Det kräver en annan förklaring.
Den bibliskt grundade slutsatsen är följande: Gud skapade människan till sin avbild, som den enda biologiska varelsen med förmåga att på ett reflekterande sätt kunna njuta av skönheten för dess egen skull. Just därför skapade Han en värld präglad av skönhet i det lilla och i det stora, där varje livsform programmerades till att uttrycka olika dimensioner av sinnrikhet och skönhet på nivåer som är möjliga att uppfattas och uppskattas av dig och mig. Detta oavsett om skönhetens ”avsändare” är en kaprifol, kiselalg, citronfjäril eller paradisfågel165. Eller ett glimrande mineral, en röd solnedgång eller en stjärnhimmel. Det är för vår skull!
↩ Tillbaka till innehållsförteckningen
![]()
Varför uppstod modern vetenskap?
Det ett ovedersägligt historiskt faktum att den moderna vetenskapen uppstod i det kristna Europa och inte i Indien, Kina, Grekland eller i den muslimska världen. Självklart gjordes upptäckter och uppfinningar även på dessa platser, men där uppstod likväl ingen organiserad vetenskap som kan jämföras med den som uppstod i väst och som idag bedrivs i hela världen. Vad beror det på?
Den frågan har diskuterats flitigt under åren, och det finns ett svar: Bibeln. Följande principer som alla har sina rötter i den bibliska uppenbarelsen anses ha bidragit till den moderna vetenskapens födelse:
1. Gud, skaparen, är en rationell varelse
Modern vetenskap vilar på grundförutsättningen att universum är begripligt och att det därför går att studera systematiskt. Det är inte så självklart som det låter. Rent historiskt vilar idén om en begriplig, lagstyrd och därmed förutsägbar värld på att den skapats av en personlig, intelligent, oföränderlig, pålitlig, lagstiftande Gud. Bibelns Gud uppenbarar sig som en rationell varelse som själv inte utgör någon del av skapelsen, vilket man däremot föreställde sig inom de österländska, panteistiska religionerna.
2. Människans uppdrag att råda över den övriga skapelsen
Enligt 1 Mos 1:28 fick människan uppdraget att råda över jorden: ”Gud välsignade dem och sa: ’Var fruktsamma och föröka er, uppfyll jorden och lägg den under er! Härska över fiskarna, fåglarna och alla djur som rör sig på jorden’.” Det uppdraget tolkades bland annat som en uppmaning till, ja till och med en befallning, att utforska skapelsen.
Ett sådant bemyndigande saknade grund inom ramen för panteismen. Snarare låg det för en panteist nära till hands att anse det vanvördigt att experimentera med den fysiska världen eftersom det i viss mening var som att experimentera med Gud själv. Inte heller antikens filosofer var särskilt intresserade av att utveckla metoder för att observera och upptäcka världen eftersom man ansåg att universum var outgrundligt.
3. Människans medvetenhet om sin fallna natur
En annan princip hämtad från Bibelns början som var en viktig pusselbit för att lägga grunden för den moderna vetenskapen finner vi i 1 Mos 3, berättelsen om syndafallet. En konsekvens av det blev en distans mellan människan och hennes skapare. Konsekvensen när människan blev avskuren från den ursprungliga intima andliga gemenskapen med honom blev att hon förvandlades till en varelse i behov av ständig tillrättavisning. Denna ofullkomlighetsprincip blev den idémässiga grundvalen för den vetenskapliga metoden, som går ut på att forskare formulerar begåvade gissningar – hypoteser – om hur världen fungerar. Dessa framställs inte som absoluta sanningar utan som preliminära modeller som sedan testas mot verkligheten. Den forskare som presenterar en hypotes hoppas att den ska stämma, men måste alltid vara beredd på att egna eller andras experiment kan motbevisa den och att en konkurrerande teori kan visa sig ha en bättre förklaringskraft. Den bärande principen är alltså en fallen och ofullkomlig människa i behov av korrigering på jakt efter den förlorade sanningen.
4. Sanning existerar
Det kanske låter självklart för somliga, men för många i vår postmoderna tid upplevs det snarast reaktionärt att hävda att sanning existerar. Men Bibeln är tydlig – det finns absoluta sanningar. Guds ord är sanning, och Jesus Kristus är sanningen själv personifierad. Allt han sade var absolut sant. Att det finns en sann och objektiv verklighet som vi kan lära känna genom att studera världen med den vetenskapliga metoden är en viktig målbild för varje forskare. Det faktum att vi genom forskningen inte når fram till den fulla sanningen är en helt annan sak. Det innebär inte att verkligheten eller sanningen som sådan är relativ eller subjektiv, att någonting är ”sant för mig men inte för dig” som blivit en postmodern devis. Sådana resonemang är farliga när de smyger sig på. Det går inte att bygga vetenskap på en sådan filosofi. I stället handlar det om att förhålla sig ödmjukt eftersom vår kunskap här i tiden är bristfällig (1 Kor 13:12).
5. Guds bud
Men det stoppar inte där. Modern vetenskap vilar på ännu en biblisk princip. Den är att Gud är en moralisk Gud som har moraliska anspråk på de människor han skapat till sin egen avbild. Utifrån en biblisk moral förväntas även forskaren agera öppet och sanningsenligt när de experimentella resultaten presenteras. Sanning och öppenhet är ingenting man kan ta för givet utifrån en ateistisk syn på vårt ursprung. I en sådan världsåskådning finns det ingen övergripande värdering som skulle göra sanning mer eftersträvansvärd eller berömvärd än lögn. Det innebär naturligtvis inte att ateister för den skull är lögnaktiga, bara att en dygd som sanningsenlighet inte kan motiveras utifrån ateistisk filosofi. Utifrån sådan går det lika bra att argumentera för att det är moraliskt att ljuga om ändamålet är gott för en själv eller andra. En sådan inställning vore givetvis förödande för vetenskapen om den fick fritt spelrum.
Tro kontra framgång under den vetenskapliga revolutionen
Den amerikanske religionssociologen Rodney Stark166 gjorde en genomgång av betydelsefulla vetenskapsmän under perioden 1543 – 1680, den period som vanligen brukar förknippas med ”den vetenskapliga revolutionen”. Av de 52 högst rankade fanns en skeptiker (Edmund Halley) och en panteist (Paracelsus). De resterande 50 var kristna, varav åtminstone 30 kan räknas som ”hängivna”, definierade av den religiösa passion med vilken de formulerade sig. Det är enligt Stark först vid tiden för Darwin som ateister åstadkom någonting att tala om inom vetenskapen (att Darwin dessvärre hade fel i det mesta är en annan femma).
Kan Starks resultat förklaras med att kristendomen var en så självklar utgångspunkt att de tidiga vetenskapsmännen inte visste bättre eller inte kände till några alternativa synsätt? Nej menar Stark – både panteism, deism och ateism och andra religiösa och filosofiska föreställningar som utmanade kristen teism var välbekanta för den tidens lärda, men man höll ändå fast vid kristendomen och argumenterade mot de alternativa synsätten därför att man ansåg att de inte höll måttet.
↩ Tillbaka till slutet av kapitel 7
↩ Tillbaka till innehållsförteckningen
![]()
Litteraturtips
Vid sidan av de böcker som redan omnämnts vill jag särskilt rekommendera:
• Ultimate Engineering: an Engineer Investigates the Biomechanics of the Human Body.167 En alldeles nyutgiven bok av Stuart Burgess, professor i ingenjörsdesign vid universitetet i Bristol.
![]()
![]()
![]()
Mars 2026
Göran Schmidt
Slutnoter
- Detta med tanke på tidigare diskussioner med Lars Johan Erkell. Mina meningsutbyten med honom finns dokumenterade på min webbplats med början på https://www.gschmidt.se/Skapelsefragan/Artiklar_Debatter/2014-2015/Bakgrunden/Bakgrunden.html ↩
- Metodologisk naturalism utgår från att alla fenomen uteslutande har naturliga (inomvärldsliga) förklaringar och att all naturvetenskap med nödvändighet måste arbeta som om Gud eller en andlig dimension inte existerar. Den är, som Erkell konstaterar, agnostisk. ↩
- Metafysisk naturalism kallas också filosofisk eller ontologisk naturalism och hävdar att inget existerar bortom det som vetenskapen kan utforska. Den är därmed ateistisk. ↩
- ”Evolution is a fact” (R. Dawkins i The Greatest Show on Earth: The Evidence for Evolution (2009). ”Well, it [evolution] is not a theory. Evolution is as solid a historical fact as you could conceive … What is a theory is wether natural selection is the mechanism…” David Attenborough, David Attenborough on Charles Darwin – Times Online av Damian Whitworth (2009). ”nearly all biologists acknowledge that evolution is a fact” Neil Campbell (i läroboken Biology 1990). ↩
- Det här anknyter till vad E. troligen menar när han skriver att ”fakta är teoriberoende”. (s 19) ↩
- Det låter på Erkell som att det bara är kreationister som gör uppdelningen av vetenskaperna i historiska och empiriska, men det stämmer inte. Det har diskuterats av William Whewell i hans bok History of the Inductive Sciences (1840), av Stephen Jay Gould i artikeln Evolution and the Triumph of Homology i American Scientist 74 (1986) s 60-69. ↩
- https://appearedtoblogly.wordpress.com/wp-content/uploads/2011/05/paley-william-natural-theology.pdf ↩
- Dels i avsnittet om ”Arrival of the fittest” på s 60 och dels i hans vederläggning av det kombinatoriska argumentet på s 109. ↩
- Simpson, G. G, The Meaning of Evolution, New Haven, CT: Yale University Press, (1967) s 344. ↩
- Parasiter kan ju många gånger tillbakabilda organ för att de inte längre behövs, som till exempel hannarna hos djuphavsmarulken. ↩
- Människan har normalt 46 kromosomer i sina celler varav 23 har kommit från den ena föräldern och lika många från den andra. Av dessa är ett par könsbestämmande (X och Y) och de resterande 22 paren kallas homologa. De kan anordnas parvis efter storlek. Rekombination sker mellan de två kromosomerna i ett sådant par. ↩
- Källa: https://pubmed.ncbi.nlm.nih.gov/11158603/ ↩
- Närmare bestämt i nr 1-2022 på sidan 36: https://genesis.nu/magasin/arkiv/genesis-2022-1/ ↩
- Med undantag från plasmaceller i vårt immunförsvar som aktivt använder sig av något som påminner om slumpgeneratorer för att kunna skapa så många olika antikroppar som möjligt. ↩
- Stone, Marcia, ”For microbes, Devolution is Evolution”, Bioscience 64 nr 10 (2014) 956. https://academic.oup.com/bioscience/article-abstract/64/10/956/1782438?redirectedFrom=fulltext. ↩
- Till exempel generna för tillverkning av flagellmotorn. Att kunna röra sig snabbt framåt är inte prio ett när man lever i en kolv med näringslösning. ↩
- Med hjälp av plasmider, så kallad horisontell genöverföring. ↩
- https://www.gschmidt.se/Skapelsefragan/Artiklar_Debatter/2014-2015/Kan_proteiner_evolvera/Kan_proteiner_evolvera_G1.pdf ↩
- Naturligtvis även allt annat levande, men det utgår jag från att du som läsare förstår. ↩
- Sanford, John C., Genetic Entropy and the Mystery of the Genome, https://www.amazon.se/Genetic-Entropy-Mystery-Genome-Sanford/dp/0981631606/ref=sr_1_1?crid=R07XYY8ZN8XE&dib=eyJ2IjoiMSJ9.ueEMuV0-N5YfCsKxJdFc3OGuUXMHBbrMzE8Vwocqwl4.jdVtpAM8-7BDz9Kr7WxBFPuoTfWTNeILnNuS5H1qjUw&dib_tag=se&keywords=genetic+entropy+-+john+sanford&qid=1769979609&sprefix=genetic+entropy%2Caps%2C181&sr=8-1. ↩
- https://www.sciencedirect.com/science/article/pii/S0022519385701671. Problemet blir förstås hanterbart om man som vi kreationister har ett hundradelen så långt tidsperspektiv på människans historia än Kondrashov. Det är bara I ett evolutionärt tidsperspektiv som problemen blir så dramatiska. Det räcker gott och väl med de miljontals genetiska defekter som mutationerna åsamkat och fortsätter att åsamka vårt genom sedan ett antal tusen år. ↩
- Skriv gärna in ”redundancy engineering” i din sökmotor och läs om det på engelskspråkiga Wikipedia. ↩
- Till exempel genom överkorsning under meiosen (könscellsbildningen) ↩
- https://www.gschmidt.se/Skapelsefragan/Artiklar/Artbildningsmekanismer%20-%20Grundl%C3%A4ggande%20mendelsk%20genetik%20-%20Genesis%203-2024%20s%2021-24.pdf ↩
- Långnosade hundraser som schäfer, blodhund och labrador (och givetvis varg och räv) har avsevärt mycket större områden med luktreceptorer vilket ger dem deras fantastiska luktsinne. ↩
- Som biblisk kreationist menar man att predation, åtminstone bland komplexa organismer, är sekundärt i skapelsen som en konsekvens av syndafallet och ingick inte i de ursprungliga ekosystemen. ↩
- Gould, S.J., Evolution’s erratic pace. Natural History 86(5):14, 1977. ↩
- För den som är bekant med begreppen homozygoti och heterozygoti från gymnasiegenetiken kan det förklaras med att inavelseffekter leder till att andelen homozygoti ökar på bekostnad av heterozygoti. Det är samma fenomen som i det nämnda hundexemplet under rubriken Selektion ovan. ↩
- Filmen kan avnjutas på https://youtu.be/2HBRTIXLCxE E. uppskattar den inte. ↩
- eller möjligen vet om men inte har något svar på, men jag tror inte det. ↩
- Närmare bestämt DNA-metylering. ↩
- Det här gäller områden som normalt är rika på C och G, så kallade CpG-öar och som har viktiga reglerande funktioner i cellen. Läs gärna mer i A Creation Science Perspective on the Origin of Diversity in Biological Life PROCEEDINGS OF THE FIRST NORDIC CREATION RESEARCH CONFERENCE, STH Academic, 2026 kap 2 s 180 ↩
- Om den här processen skulle fortsätta tillräckligt länge skulle DNA-molekylerna till slut bara innehålla A och T och inget G och C. Då skulle DNA:s förmåga att bära information för länge sedan ha upphört. Det råder alltså ingen tvekan om att de tendenser i denna riktning som observeras verkar åt fel håll för evolutionen. ↩
- Exempelvis har alla de olika arterna av ciklider i de tre stora afrikanska sjöarna i princip identiskt DNA. Det är epigenetiken och inte DNA-sekvensen som är orsaken (se Proceedings s 162) (se not 32 ovan) ↩
- https://www.gschmidt.se/Skapelsefragan/Artiklar_Debatter/2014-2015/Biologisk_information/Biologisk_information2.html#xyz ↩
- Det fanns en tidigare term med samma betydelse, nämligen preadaption (”anpassning i förväg”), men man övergav det eftersom det gav intrycket av att evolutionen hade förutseende. ↩
- Ordspråket förmedlar tanken att det är omöjligt att få något utan att ge något i gengäld. ↩
- Se nr 1-2022 på https://genesis.nu/magasin/arkiv/genesis-2022-1/. Artikeln finns även fristående för nedladdning på min egen webbplats: https://www.gschmidt.se/Skapelsefragan/Artiklar/Tredje%20v%C3%A4gens%20evolutionsbiologi%20-%209%20Endosymbiontteorin%20-%20Genesis%201-2022%20s%2046-49.pdf. Se även den mer omfattande artikeln om endosymbiontteorin av Jerry Bergman på https://creation.com/en/articles/research-overturned-endosymbiosis/. ↩
- Källa: Werner, Carl, ”Living fossils”, New Leaf Press, 2008, s 169-182, 263. ↩
- Flew är den övertygade ateisten som på äldre dag kom till slutsatsen att molekylärbiologin vittnade om en skapare. Hans självbiografi: There is no a God rekommenderas varmt. Bokens fullständiga titel är: There is a God - How the World's Most Notorious Atheist Changed His Mind, HarperOne 2008, ISBN: 9780061335303. ↩
- För ryggradsdjuren räknar man inom baraminologin (den bibelbaserade systematiken) att gränsen för skapade slag oftast går mellan de olika famiE.rna (hunddjur, kattdjur etc). För andra djurstammar kan nivån vara annorlunda. ↩
- https://www.youtube.com/watch?v=AD4sYIxBluE&list=PLNkVOx7YbWiDABv5nB3IhPNi1g14SPHO_ ↩
- Hössjer O., Lampa S., Schmidt G., ”A Creation Science Perspective on the Origin of Diversity in Biological Life”, STH Academic, 2026, kap 2, s 78-121 ↩
- https://en.wikipedia.org/wiki/World_population#/media/File:Population_curve.svg ↩
- 1. På Noas ark fanns ”anfäderna till de djur som finns idag”. Mer korrekt är ”anfäderna till de familjer av större, landlevande utdöda och nutida ryggradsdjur som finns idag motsvarande (inalles ca 2000 individer). 2. ”Man sätter … sin tolkning av Bibelns skapelseberättelser framför den etablerade vetenskapliga synen.” Men Genesis anser inte att Bibeln har fler än en skapelseberättelse. ↩
- Se https://genesis.nu/faq/vad-anser-ni-talar-mest-skapelseteorin/. ↩
- Se Genesis temanummer om teistisk evolution: https://genesis.nu/magasin/arkiv/genesis-2018-2/ ↩
- Naturligtvis förnekas detta av evolutionister, i synnerhet i polemik mot kreationister, men förhållandet är väl förankrat även i den sekulära litteraturen och av framträdande evolutionsbiologer, som till exempel: Stephen J. Gould: ”Den extrema sällsyntheten av övergångsformer i fossilens arkiv är och förblir paleontologins affärshemlighet.” (Evolution’s erratic pace. Natural History 86(5):14, 1977) Hickman C.P. et al: ”Många arter förblir praktiskt taget oförändrade under miljontals år varefter de försvinner och ersätts av en tämligen annorlunda men besläktad form. Dessutom uppträder de flesta högre kategorier av djur abrupt i fossilarkivet, fullt utvecklade, och utan några hittills upptäckta fossil som bildar en övergång från deras föräldragrupp.” (Hickman C. P., Roberts, L. S., Hickman F. M. Integrated Principles of Zoology (1988, 8:e upplagan, s 866) och Ernst Mayer: ”Nya arter uppträder vanligtvis plötsligt i det fossila arkivet utan att vara förbundna med sina förfäder genom en serie av mellanformer” (What Evolution Is, Basic Books, 2001, s 189), och ↩
- https://scienceandculture.com/2021/06/recognizing-design-by-a-purposeful-arrangement-of-parts/ ↩
- Att den här texten skulle vara AI-genererad är förstås en teoretisk möjlighet, men det hade varit ett utmärkt tillfälle att testa den typ av designdetektering som vi talar om för att räta ut det frågetecknet. ↩
- Dembski, William A., Intelligent Design – The Bridge Between Science and Theology, InterVarsity Press (1999). ↩
- S.E.T.I. står för Search for ExtraTerrestrial Intelligence, alltså sökandet efter utomjordisk intelligens. ↩
- Boken filmatiserades 1997. ↩
- Primtal är den talserie som består av alla tal större än 1 som inte är jämnt delbara med något annat tal än 1 och sig självt, det vill säga 2,3,5,7,11,13,17,19,23 o s v. ↩
- https://www.astro.uu.se/~ez/hephaistos/hephaistos.html ↩
- Till exempel i Ps 8 och Rom 1. ↩
- Det är, förmodar jag, också den gud som de religiösa personer föredrar som E. låtit förhandsläsa boken. De namnges på s 8 i bokens förord och är prästerna i Svenska Kyrkan: Fredrik Ivarsson, Margaretha Josefsson, Sara Wrige och Jonas Eek, samt judiske rabbinen Peter Borenstein. ↩
(Tillbaka till förordet: ↩)
- Halmdocka eller ”straw man” är när man i stället för det verkliga argumentet konstruerar ett liknande argument och angriper det i stället, på grund av att man inte kommer åt att angripa det verkliga. Det är ett klassiskt logiskt felslut. ↩
- Eukaryoter är organismer som har huvudparten av sitt DNA samlat i en cellkärna. Summan av allt DNA kallas organismens genom (uttalas jenåm, med betoning på sista stavelsen). ↩
- Om du läst Erkells bok så finns där ett appendix som utgör den vetenskapligt korrekta delen av boken. Den är nämligen rent deskriptiv och fri från E.s egna värderingar. ↩
- Uttalas ”splajseosåmen, med betoning på å:t. ↩
- Antalet olika celltyper i en människa brukar uppskattas till ungefär 250. ↩
- Mattick, J. Amaral, P. RNA the Epicenter of Genetic Information, Taylor&Francis 2023 s. 157. ↩
- Kellis M. et al., Defining functional DNA elements in the human genome, april 2014, doi: 10.1073/pnas.1318948111. ↩
- Mattick John, Amaral Paulo, RNA, the Epicenter of Genetic Information (2022) https://www.taylorfrancis.com/books/oa-mono/10.1201/9781003109242/rna-epicenter-genetic-information-john-mattick-paulo-amaral ↩
- Se https://genesis.nu/magasin/arkiv/genesis-2025-2/. Numret blir tillgängligt för icke-prenumeranter 2026-06-01, men går att beställa i pappersform. En artikel från numret som ger grunddragen i systemargumentet finns även på min webbplats: https://www.gschmidt.se/Skapelsefragan/Artiklar/Abiogenes%20-%20Om%20system%20och%20design%20-%20Genesis%202-2021%20s%2040-42.pdf ↩
- Till exempel andningssystem, cirkulationssystem och nervsystem m fl som alla är beroende av varandra i en levande organism). ↩
- Bland annat en evolutionär spekulativ berättelse om hur en så kallad injektisom (proteinspruta som vissa bakterier har) skulle ha omvandlats till en flagellmotor, den inombordsmotor som ger vissa arter av en särskild form av rörelseförmåga. ↩
- Stephen Meyer skriver i sin bok Signaturen i Cellen frågan: ”Om utsagan ’Jupiter består huvudsakligen av metangas’ är en vetenskaplig utsaga bör även utsagan ’Jupiter består inte huvudsakligen av metangas’ vara vetenskaplig. Negationen av en utsaga gör den inte till en annan sorts utsaga. På samma sätt är inte utsagan ’Intrycket av design inom biologin är inte ett resultat av reell design’ och påståendet ’intrycket av design i biologin är ett resultat av design’ inte två olika slag av utsagor; de utgör två olika svar på samma fråga, en fråga som länge varit en del av evolutionsbiologi och historisk vetenskap. Om den ena av dessa utsagor är vetenskaplig förefaller det som om även den andra är det.” (s 411) ↩
- Behe M. The Edge of Evolution s 57. ↩
- Sannolikheten att till exempel slå två tärningar och få sexor på båda är 1/6 x 1/6 = 1/36, så man får kasta tärningarna i genomsnitt 36 gånger innan man kan räkna med att det händer. ↩
- Det här framgår tydligt av figur 3 i https://www.pnas.org/doi/10.1073/pnas.1322965111 ↩
- https://scienceandculture.com/2014/07/a_key_inference/ ↩
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1173140/ ↩
- Sådant är urvalet nämligen bra på, att finkalibrera existerande egenskaper. Eller som de Vries sade: survival of the fittest, men inte arrival of the fittest. (se Selektion s 16). ↩
- Den bakteriepopulation man utgick från vid försökets start bestod till 99% av en muterad form av E. coli där genen malT var utslagen av en mutation för att förhindra angrepp av lambdafager som utnyttjade det motsvarande cellyteproteinet som ”landningsbana” innan det kunde angripa cellen. Det innebar att 99% av bakterierna i populationen inte längre kunde omsätta sockerarten maltos för att de saknade transportprotein för detta. Bara 1% av dem hade intakta proteiner som kunde transportera in maltos i cellerna, och som därmed kunde tjäna som landningsbana för fagerna. ↩
- https://scienceandculture.com/2012/01/more_from_lensk/ ↩
- I praktiken 21, men det kan den intresserade fråga ChatGPT om. ↩
- Annars kallas den rätt och slätt polypeptid, eftersom det alltid är en speciell kemisk koppling mellan aminosyrorna som kallas peptidbindning. ↩
- Ifall någon skulle undra hur det då kan komma sig att det bildas proteiner inuti cellerna precis hela tiden, så beror det på att de platser där de tillverkas (vid de så kallade ribosomerna) utgörs av vattenavstötande fickor i ribosomproteinet (ribosomerna består av en kombination av RNA (rRNA) och protein). ↩
- Det är inte utrymme att utveckla det här, men faktum är att inte ens den ynka rekvisitan kan E. ta för given. Ännu så länge finns det inte ett enda livets-uppkomst-experiment under realistiska förhållanden som genererat vare sig användbara (nämligen i icke-racemiska blandningar för den som vill googla) aminosyror eller ens en enda nukleotid (John Walton, The Times Litterary Supplement 5567 (letter to the editor): 6 (11/12 2009) ↩
- Jag valde ordet ”blygsam” trots att det inte är vetenskapligt korrekt. Det är som vi ska se empiriskt välbelagt att det är en väldig informationsmängd som krävs för fortplantning. ↩
- Eller möjligen en AI-assistent. Men denna är i sin tur designad av varelser med de egenskaperna och inlärd på väldiga mängder information producerad av sådana. ↩
- Dessutom gör Dawkins i sin evolutionssimulering ett antal mycket grova och orealistiska förenklingar:en naturlig mutationsfrekvens på 1/10 000 000 000 byts ut mot 1/28. organismens arvsmassa reduceras till en bråkdel (från storleksordningen 3 miljarder till 28 informationsenheter).en under naturliga förhållanden okänd men som regel mycket liten selektionschans för en positiv mutation byts ut mot en hundraprocentig sådan.varje selektion ökar textsträngens informationsinnehåll med en enhet. Detta är i verkligheten ytterst osannolikt.Det är bland annat alla dessa förenklingar som gör att Dawkins metod konvergerar. Utan dessa förenklingar är det enkelt att visa att Dawkins dator skulle behöva mer tid på sig än de 13,8 miljarder år som Big Bang-teorin medger. ↩
- Matematikern David Berlinsky kallar detta ”kombinatorisk inflation” i sin bok The Deniable Darwin från 1996. ↩
- I många proteiner, speciellt hos mer komplexa organismer som människan är det vanligt att proteiner innehåller avsnitt som inte är stabila, utan ”sladdriga”, men det ingår då som en viktig del i deras totala funktion, så det säger inte att aminosyrorna i de avsnitten kan bytas ut hur som helst. ↩
- Axe, Estimating the Prevalence of Protein Sequences Adopting Functional Protein Folds. ↩
- En proteinfamilj består av ett antal proteiner med liknande sekvens av aminosyror och därför likartad tredimensionell struktur. Det finns tiotusentals dokumenterade sådana. ↩
- Durston et al., Measuring the Functional Sequence Complexity of Proteins, Reidhaar-Olson och Sauer, Functionally acceptable Solutions in Two Alpha-Helical Regions of Lambda Repressor, Taylor et al., Searching sequence Space for Protein Catalysts, Yockey, A Calculation of the Probability of Spontaneous Biogenesis by Information Theory. ↩
- Till exempel i artikeln The Roles of Mutation, inbreedning, crossbreeding, and selection in evolution från 1932. ↩
- https://www.jcvi.org/media-center/first-minimal-synthetic-bacterial-cell-designed-and-constructed-scientists-venter?utm_source=chatgpt.com ↩
- En nackdel med Shannons informationsbegrepp är att symbolföljden (1) faktiskt innehåller mer information än nummer (3). Nummer (1) är helt slumpmässig, medan nummer (3) präglas av viss regelbundenhet (t ex att en vokal i svenska språket oftast följs av en konsonant) som gör att den är något mindre osannolik, och därmed bär mindre information. ↩
- Exempel är Stephen Meyer, William Dembski och Werner Gitt. ↩
- Komplex i bemärkelsen att den är väldigt osannolik. Chansen att en slumpgenerator eller flock schimpanser skulle upprepa den är så liten att det skulle förväntas ta oräkneliga miljarder år innan den skulle uppträda igen. ↩
- Virus räknas inte som en livsform på grund av dess oförmåga till fortplantning utan tillgång till en levande cell. Även många virus innehåller DNA, även om merparten antas innehålla RNA som informationsbärare. ↩
- Huruvida de återstående 98,5 procenten av människans arvsmassa är funktionella eller inte återkommer vi till längre fram. ↩
- Här skulle E. möjligen protestera och hävda att det är helt uppenbart att evolutionen gjort det med hänvisning till all den information som de facto finns i jordens alla varelser, men det vore naturligtvis att förutsätta det (evolutionen) som skulle bevisas, alltså ett uppenbart cirkelresonemang. ↩
- Hazen et. al. i Proceedings of the National Academy of Sciences (2007): ”Functional information and the emergence of biocomplexity”, https://pmc.ncbi.nlm.nih.gov/articles/PMC1876432/. ↩
- Människans haploida arvsmassa består av 3,2 miljarder kvävebaser (3,2 Gb), där varje bas/bokstav motsvarar två bitar Shannoninformation. Totalt skulle det, lågt räknat, ge ett informationsinnehåll på 6,4 Gbit (eller 0,8 Gbyte) i varje människocell. ↩
- Det bör tilläggas att detta scenario påstås ha skett utan någon verkande selektion. Scenariot är orimligt med selektion och närmast absurt utan. Föreställningen hålls vid liv huvudsakligen tack vare att duplicering är den enda kända process som kan öka storleken på en arvsmassa. ↩
- E. presenterar inga experimentella resultat som stödjer hans uppfattning. Men i en tidigare debatt med mig nämner han fyra exempel: De handlar om maltasenzymer, glukokortikoidreceptorn, protonpumpen och citratätande bakterier. Meningsutbytet finns dokumenterad på min webbsida på https://www.gschmidt.se/Skapelsefragan/Artiklar_Debatter/2014-2015/Biologisk_information/Biologisk_information.html ↩
- Det är i grunden samma anledning som att det är i stort sett omöjligt att ändra på en hemsidas struktur när den väl har sjösatts. Att ändra grundstrukturen innebär att referenser tappar sina mål och leder till dialogrutan ”webbplatsen kan inte nås” (401-fel). ↩
- En av djurgrupperna ligger helt fel i diagrammet. Snabelsäcksmaskar är typiska för den kambriska faunan. ↩
- Foote, Sampling, Taxonomic Description and Our Evolving Knowledge of Morphological Diversity, s 181 ↩
- Se till exempel de genregulatoriska illustrationerna i https://www.pnas.org/doi/10.1073/pnas.0711220105 ↩
- Nu är Meyer inte ungjordskreationist (ännu) men även räknat i geologisk tid är perioden kortvarig. ↩
- 10 000 är den populationsstorlek som evolutionsbiologer anser förelåg när människan utvecklades. ↩
- https://www.sciencedirect.com/science/article/abs/pii/S0040580997913587?via%3Dihub ↩
- https://pmc.ncbi.nlm.nih.gov/articles/PMC1950772/?utm_source=chatgpt.com ↩
- https://academic.oup.com/genetics/article-abstract/180/3/1501/6063886?redirectedFrom=fulltext och https://link.springer.com/article/10.1186/s12976-015-0016-z ↩
- https://link.springer.com/chapter/10.1007/978-3-030-02825-1_12 och https://www.sciencedirect.com/science/article/pii/S0022519321000795?via%3Dihub ↩
- Till exempel Barnard, F.M., and A. Maxwell. 2001. Interaction between DNA gyrase and quinolones: effects of alanine mutations at GryA subunit residues Ser83 and Asp87, Antimicrobial Agents and Chemotherapy 45:1994–2000. ↩
- Graden av specificitet är direkt kopplat till informationsinnehållet. Om jag ber min son gå en i en butik och köpa te, så behöver jag inte använda särskilt många ord (informationsenheter). Det räcker att jag säger till honom: - Köp te! (ty han är en lydig 37- åring ;). Om jag däremot vill ha tepåsar med Earl Grey av märket Twinings i 250-gramsförpackning är min begäran mer specifik och kräver därför en betydligt längre specifikation – mer instruktioner = information. ↩
- Det är för övrigt samma sak med begreppet ”anpassad” inom biologin. Att ett djur eller en växt är anpassad till sin miljö behöver inte alls bero på mutationer och selektion, det är ett minst lika möjligt alternativ att de är anpassade därför att Gud i begynnelsen anpassade dem till att leva i en föränderlig värld och därför utrustade dem med allehanda system för just det syftet, som till exempel epigenetiska mekanismer. ↩
- Fumagalli, R. The Universe’s Fine-Tuning Does Call for Explanation. J Gen Philos Sci (2025). https://doi.org/10.1007/s10838-025-09734-8. ↩
- Antropocentrisk betyder att människan sätts i centrum och ses som måttstock för hur världen förstås och värderas. Ett antropocentriskt synsätt utgår från att natur, djur och miljö främst har betydelse utifrån hur de påverkar människan. (Källa: ChatGPT) ↩
- Det här är anledningen till att antalet olika proteiner i en människa är ungefär 300 gånger större än antalet proteinkodande gener. ↩
- Viskositet är hur trögflytande en vätska är. Sirap har hög viskositet i förhållande till vatten. ↩
- Ämnen med hög viskositet som sirap ger låga Reynoldstal, medan lättflytande vätskor och i synnerhet gaser ger mycket höga värden. Storleken på talet avgör om strömningen blir laminär (rätlinjig) eller turbulent (med virvelbildning). Talet saknar enhet men används för konstruktion av flygplan, fartyg med mera. Molekylära motorer arbetar vid extremt låga reynoldstal. ↩
- Den läsare som är intresserad av den här sortens nanoteknologi rekommenderas för övrigt en videoserie av Dr James Tour på Youtube: https://www.youtube.com/playlist?list=PLILWudw_84t1d-Hxvf5VnJxCmi4X3lxCR ↩
- https://www.gschmidt.se/Skapelsefragan/Artiklar_Debatter/2014-2015/Molekylara_maskiner/Molekylara_maskiner.html ↩
- https://www.gschmidt.se/Skapelsefragan/Artiklar_Debatter/2014-2015/Sammanfattning/Sammanfattning.html ↩
- Se till exempel https://www.gschmidt.se/Skapelsefragan/Artiklar/Abiogenes%20-%20Vad%20%C3%A4r%20liv%20-%20Genesis%201-2019%20s%2010-13.pdf och https://www.gschmidt.se/Skapelsefragan/Artiklar/Tredje%20v%C3%A4gens%20evolutionsbiologi%20-%201%20Sj%C3%A4lvorganisation%20-%20Genesis%201-2022%20s%2020-23.pdf ↩
- https://www.gschmidt.se/Skapelsefragan/Artiklar/Virus%20-%20Genesis%202-2022%20s%2046-49.pdf ↩
- https://liu.diva-portal.org/smash/record.jsf?pid=diva2%3A617381&dswid=160 ↩
- Att jordens livsformer faktiskt är designade top-down bekräftas av att den bara 20 år gamla biologivetenskapen systembiologi haft sådana framgångar. Den utgår nämligen just från ett sådant perspektiv på livet. ↩
- Det är bekvämt för evolutionsbiologer som därför inte tycker sig behöva förklara det märkliga förhållandet att obesläktade varelser har samma ordning på tusentals ”bokstäver” i sitt DNA. Det finns förklaringar för bakterier, för de kan utbyta DNA genom så kallade plasmider. Det kallas horisontell genöverföring (Horisontal Gene Transfer, HGT). Det saknas helt evidens för att det kan ske hos ryggradsdjur. ↩
- Läs gärna mer om det här i Genesis temanummer om System: https://genesis.nu/magasin/arkiv/genesis-2025-2/ när det blir publikt 1/6 2026, eller beställ via föreningens hemsida. ↩
- Crick var en av DNA-molekylens upptäckare. ↩
- A Devil's Chaplain: Reflections on Hope, Lies, Science and Love, s. 79, 2001. ↩
- The Blind Watchmaker, s 1, 1987 ↩
- What Mad Pursuit: A Personal View of Scientific Discovery, 1988 ↩
- Deborah Kelemen, professor i psykologi vid universitetet i Boston konstaterar att: "även om avancerad vetenskaplig träning kan reducera accepterandet av vetenskapligt felaktiga teleologiska (målinriktade – min anm.)förklaringar så kan den inte utplåna en envis, tidigt uppträdande, mänsklig tendens att finna ändamålsenlighet i naturen.", se Art Jahnke, The Natural Design Default: Why Even the Best-Trained Scientists Should Think Twice, Bostonia, W/S 2013. https://www.bu.edu/bostonia/winter-spring13/the-natural-design-default/ , https://www.bu.edu/bostonia/winter-spring13/the-natural-design-default/ ↩
- Alison Gopnik, professor i psykologi vid Berkeleyuniversitetet, See Jane evolve: Picture Books Explain Darwin, Mind and Matter, Wall Street Journal, 18 april 2014. https://www.wsj.com/articles/SB10001424052702304311204579505574046805070 ↩
- https://www.gschmidt.se/Manadens_reflektion/Reflektioner/2016/Augusti/Augusti_2016.html ↩
- Vilket givetvis inte utesluter att fenomen som stenar och katter som sådana skulle vara designade. ↩
- Det finns evidens för att både resistensmutationer mot malaria och sömnsjuka (Trypanosomas) faktiskt uppstår mer frekvent hos befolkningar i malaria- och sömnsjuketäta områden än den rent statistiskt borde göra. (se t ex: https://www.biorxiv.org/content/10.1101/2021.05.24.443729v1 och https://pubmed.ncbi.nlm.nih.gov/40854136/ . Se även: https://phys.org/news/2025-09-mutations-evolution-genome-random.htm l. För mer information om riktade mutationer - se s 54 och 164 i A Creation Science Perspective on the Origin of Diversity in Biological Life PROCEEDINGS OF THE FIRST NORDIC CREATION RESEARCH CONFERENCE, STH Academic, 2026. ↩
- https://www.gschmidt.se/Skapelsefragan/Artiklar/Tredje%20v%C3%A4gens%20evolutionsbiologi%20-%201%20Sj%C3%A4lvorganisation%20-%20Genesis%201-2022%20s%2020-23.pdf ↩
- Som för övrigt är en i mitt tycke utmärkt nyhetssajt. Du hittar den på https://scienceandculture.com/ ↩
- https://ncse.ngo/files/pub/creationism/The_Wedge_Strategy.pdf ↩
- https://roots.discovery.org/. ↩
- https://www.garynorth.com/freebooks/docs/pdf/the_changing_of_the_guard.pdf ↩
- https://christianreconstructionist.wordpress.com/wp-content/uploads/2017/12/institutes-of-biblical-law-vol-1-rushdoony.pdf ↩
- Johnson, Phillip E.,”The Wedge of Truth – Splitting the Foundations of Naturalism”, InterVarsity Press, 2000. Citatet är från s 83, översättning DeepL. ↩
- Darwin, Charles, ”The origin of species by means of natural selection, or, The preservation of favored races in the struggle for life”, Modern Library Edition, Random House Inc., 1993 (1859), s 19 (förf. övers.). ↩
- https://genesis.nu/magasin/arkiv/genesis-2021-3/ ↩
- https://www.arn.org/docs/dembski/wd_theologn.htm ↩
- Se artikeln på s 37 i magasinet Genesis 4-2024: https://genesis.nu/site/assets/files/4295/genesis-2024-4.pdf ↩
- https://www.discovery.org/m/2018/12/Parents-Guide-to-Intelligent-Design.pdf s 7. ↩
- Nagel, Thomas, The Last Word, pp. 130–131, Oxford University Press, 1997. Dr Nagel (1937– ) (professor i filosofi och juridik vid New York University). ↩
- I synnerhet om vi hade pratat om sorkar eller älgar som inte kan gå till optikern och få glasögon. ↩
- Det är mycket svårt att tillverka bara den ena spegelbilden av ett ämne i ett laboratorium. Ett bevis på det är att det gav Nobelpris i kemi 2001. De två spegelbilderna av ett ämne, t ex ett läkemedel, kan ha helt olika effekt i kroppen. Det finns tragiska exempel på det, som talidomid (”neurosedyn”). Läs om det på nätet om du vill. ↩
- En av de naturligt förekommande aminosyrorna – glycin – saknar spegelbildsform. Det beror på att den har en väteatom på den plats där det står ett R i figuren, och då har kolatomen bara tre olika atomgrupper knutna till sig. I de övriga 19 aminosyrorna sitter det på R:ets plats en större eller mindre kedja av kolatomer. ↩
- I stället för beteckningarna L/D används ibland vänster/höger eller S/R, men det är samma sak (bortsett från att aminosyran cystein i levande varelser i det nya systemet blir definierat som R i stället för S). ↩
- Det finns nämligen en teknik för att framställa rena former av de båda spegelbildsformerna, men då måste man använda sig av biologiskt bildade homokirala ämnen som ”mall”. ↩
- Det förekommer enstaka undantag, som t ex i cellväggarna på vissa bakterier. En trolig tolkning är att det är ett ”försvarsstrategi” för bakterierna, eftersom enzymer är anpassade till att bara klippa sönder kedjor med en och samma spegelbildsform. ↩
- Somliga forskare menar att det kanske fanns en situation där det råkade vara 90 procent av den ena varianten och 10 procent av den andra. Då skulle chansen vara mycket större. Men räknar man på det blir chansen ändå bara en på 5 x 1013, d v s en på 50 tusen miljarder (0,9300). Det är förstås en mycket större chans, men det handlar om en enda ensam molekyl. Och en levande varelse består inte av en enda molekyl utan av miljarders miljarder. Man kan också tillägga att även om slumpen verkligen hade lyckats åstadkomma en kedja av bara den ena spegelbilden av aminosyror, så skulle den innehålla lika mycket meningsfull information som en slumpmässig följd av versaler, d v s ingen alls. Att skapa meningsfull information på en molekylkedja med 300 aminosyrarester skulle vara nästa stora utmaning. Det vore ännu mindre sannolikt än att bilda proteinkedjor, eftersom det inte finns två alternativ av aminosyror, utan 20 (29 om det hade handlat om svenska versala bokstäver). ↩
- https://genesis.nu/magasin/arkiv/genesis-2020-2/ s 31-34. ↩
- Begreppet fitness är ett ord som används flitigt i evolutionssammanhang. Det betyder helt enkelt anpassningsgrad. En organism med hög fitness är välanpassad till sin miljö och får statistiskt sett fler ungar än sådana med lägre. ↩
- Det har nyligen visat sig att evolutionsbiologers observationer på det här området ofta varit omöjliga att återupprepa. Evolutionära förväntningar har alltför ofta tillåtits påverka resultaten: https://www.the-scientist.com/features/replication-failures-highlight-biases-in-ecology-and-evolution-science-64475 ↩
- Se t ex: Being preened to perfection is no guarantee of success, New Scientist 197(2649):16, 2008. ↩
- https://www.newscientist.com/article/dn13535-have-peacock-tails-lost-their-sexual-allure/ Takahashi, M. and others, Peahens do not prefer peacocks with more elaborate trains, Animal Behaviour 75(4):1209–1219, 2008 | doi:10.1016/j.anbehav.2007.10.004. ↩
- Det finns även forskning som visar på att hannar väljer honor efter yttre attribut. Kanske inte helt obekant inom vår egen art 😉, men det är betydligt mindre utforskat. I djurvärlden är det ju oftast hannarna som är färggrannast, även om det finns enskilda undantag (t ex blågyltan – en västkustsk läppfisk) ↩
- https://www.ted.com/talks/denis_dutton_a_darwinian_theory_of_beauty#t-3828. ↩
- Om du har något kvar av skatteåterbäringen som du inte vet hur du ska spendera, så överväg att införskaffa denna magnifika bok om lövsalsfåglar och paradisfåglar. Den kan ligga där på soffbordet som ett vittnes-bird 😉 om Skaparens kreativitet och sinne för skönhet https://www.bokus.com/bok/9780691202143/birds-of-paradise-and-bowerbirds/ ↩
- For The Glory of God: How Monotheism Led to Reformations, Science, Witch-Hunts and the End of Slavery av Rodney Stark, Princeton University Press, Princeton, 2003. En recension finns på https://answersingenesis.org/reviews/books/the-biblical-origins-of-science/ ↩
- https://www.bokus.com/bok/9781637120842/ultimate-engineering/ (kortare: bit.ly/MTBK70). Se en presentationsvideo av boken på https://youtu.be/G0rcGJbI4Yg (kortare: bit.ly/MTBK71) ↩